in wild plants and
animals in Sweden
A review of case studies from the
perspective of conservation genetics
REPORT 5786 • JAN 2008
Chromosome Copy of the chromosome Cell Cell nucleus Mitochondria Nuclear DNA Mitochondrial DNA Supportive proteins Base pairs
and animals in Sweden
A review of case studies from the perspective of conservation geneticsAnna-Carin Lundqvist
Department of Evolution Genomics and Systematics Evolutionary Functional Genomics
Uppsala University Stefan Andersson Department of Ecology Plant Ecology and Systematics
Lund University Mikael Lönn School of Life Sciences
Molecular Ecology Södertörn University College
Address: CM-Gruppen, Box 110 93, SE-161 11 Bromma, Sweden
Internet: www.naturvardsverket.se/bokhandeln
The Swedish Environmental Protection Agency
Phone: + 46 (0)8-698 10 00, Fax: + 46 (0)8-20 29 25 E-mail: natur@naturvardsverket.se
Address: Naturvårdsverket, SE-106 48 Stockholm, Sweden Internet: www.naturvardsverket.se
ISBN 978-91-620-5786-2.pdf ISSN 0282-7298 © Naturvårdsverket 2007 Tryck: CM Digitaltryck AB, Bromma 2008
Layout: Naturvårdsverket and Press Art Cover photos:
Chromosome, illustration: Anna-Carin Lundqvist. Wolf (Canis lupus) photo: Myra bildbyrå. Field fleawort (Tephroseris integrifolia) photo: Björn Widén Permission to publish Figure 2 is given by Wiley-Blackwell
(Oxford, UK) and for the remaining photos from the respective photographer or company.
Preface
This overview was initiated to get an updated report on what is known about the genetic variation of wild populations of plants and animals in Sweden. The project was part of a government commission to the Swedish Environ-mental Protection Agency to suggest an action programme for conservation of genetic variation in wild plants and animals in Sweden. This is part of the work to achieve the national environmental quality objective A rich diversity
of plant and animal life (http://miljomal.nu/english/english.php) which was
adopted by the Swedish Parliament in 2005. The overview has been written by Dr Anna-Carin Lundqvist (Department of Evolution, Genomics and Syste-matics, Uppsala University), and Associated Professors Stefan Andersson (Dept. of Plant Ecology and Systematics, Lund University) and Mikael Lönn (School of Life Sciences, Södertörn University College, Stockholm), with Seni-or Adviser Per Sjögren-Gulve (The Wildlife Management Unit of the Swedish EPA) as project leader and managing editor. The views presented in this report are those of the authors and should not be taken as those of the Swe-dish EPA.
The overview is written for managers and decision-makers working with natural resources, nature- and species-conservation, and sustainable develop-ment, at central and local government agencies, ministries, in municipalities and in non-governmental organizations. Chapter 1 explains the focus of the report and gives a brief background relating to basic population genetics. Chapter 2 describes positive outcomes associated with conserving genetic variation. Chapters 3-7 review patterns in the genetic variation of wild plants and animals in Sweden, and relevant underlying processes, on the basis of selected themes with particular relevance to biodiversity conservation. Each chapter starts with a summary, followed by a general review of the internatio-nal literature in the area and by illustrative examples, primarily using Swedish case studies. Chapter 8 then discusses important principles and analyses that need to be considered in a genetic monitoring programme, and chapter 9 pre-sents the conclusions from the overview.
The overview has been peer reviewed by senior scientists and a draft was discussed and reviewed by scientists and managers in a reference group and at a seminar. The Swedish EPA thanks all who have contributed in the process and hopes that the report will provide useful insights and examples for the work with conservation of ecosystems, species, populations and genetic varia-tion, in Europe and other parts of the world.
Stockholm, December 2007
Björn Risinger
Contents
PREFACE 3
CONTENTS 4
SUMMARY 7
1. BACKGROUND 12
1.1 The aim and focus of the report 13
1.2 Basic theory of population genetics 15
2. BENEFITS OF GENETIC DIVERSITY 17
2.1 Why should we conserve genetic variation? 17
2.2 The positive effects of genetic variation at the population,
community and ecosystem levels 19
3. LOSS OF GENETIC DIVERSITY IN SMALL POPULATIONS 23
3.1 Loss of genetic variation on the individual level 24
3.1.1 Studies of natural populations 26
3.2 Effects on fertility and viability 29
3.2.1 The effects on fertility and viability in natural populations 30 3.3 The effects of loss of genetic variation on the long term ability to adapt 32
3.3.1 Studies investigating the effects on long term adaptive abilities in natural
populations 33
4. GENETIC DIVERSITY AFTER GENE FLOW AND HYBRIDISATION 35
4.1 Hybridisation and gene flow in theory 36
4.2 Hybridisation and gene flow in natural populations 37
4.2.1 Hybrid fitness 37
4.2.2 Gene flow and hybridisation after environmental disturbance 38 4.2.3 Genetic contamination by introduced species or genotypes 38 4.2.4 Outbreeding depression and genetic assimilation 42
4.2.5 Transgenes and heterosis effects 44
4.3 Importance for the formation of species and conservation values 44 4.4 Genetic aspects of restoration and supplementation projects 45
5. GENETIC EFFECTS OF HARVESTING 48
6. GENETIC DIVERSITY AND CHANGE OF HABITAT AND CLIMATE 52
6.1 Local adaptation 52
6.1.1 Transplantation experiments 53
6.1.2 Genetic differences between environments – molecular genetic markers 54 6.1.3 Genetic differentiation between environments – quantitative characters 56
6.1.4 The possibility of future adaptations 58 6.2 Local adaptation in association with changes of distribution and climate 59
6.3 Marginal and central populations 61
7. GENETICALLY DISTINCT POPULATIONS IN SWEDEN 63
7.1 Genetic or taxonomic variation? 64
7.2 The conservation value of Scandinavian populations 66 7.2.1 Endemism in Scandinavian vascular plants 68 7.2.2 Genetic studies of terrestrial plants and animals 68 7.2.3 Genetic studies of aquatic plants and animals 72 7.3 Research progress and continued lack of knowledge 72
8. GENETIC MONITORING 75
8.1 Which taxa are in need of genetic monitoring? 76
8.2 Genetic monitoring – theory and practice 79
8.2.1 General principles of a genetic monitoring programme 79 8.2.2 Identifying populations showing negative population trends 80
8.2.3 Genetic monitoring in practice 80
9. CONCLUSIONS 83
10. ACKNOWLEDGEMENTS 87
Summary
By ratifying the Convention of Biological Diversity, Sweden has agreed to conserve the biological diversity at the ecosystem, species and genetic levels. One common assumption is that the conservation of ecosystems and habitats also conserves species, and that the conservation of species also conserves genetic diversity. However, there is a growing realization that the conserva-tion of species does not necessarily conserve the genetic diversity within speci-es. Until now, the conservation of genetic resources has attracted relatively little attention in practical nature conservation.
In recent years several scientists have argued that the conservation of bio-logical diversity should focus on preventing the disappearance of genetically distinct populations rather than to solely prevent the extinction of species. To conserve genetically distinct populations could be a better way of conserving the evolutionarily potential of species. This will also reduce the risk that spe-cies go extinct, even in a longer time perspective.
In 2006, The Swedish Environmental Protection Agency received a government commission to develop a national action plan for the conserva-tion of genetic diversity in wild plants and animals, in consultaconserva-tion with the Swedish Board of Agriculture, the Swedish Forest Agency, the Swedish Board of Fisheries and the Swedish University of Agricultural Sciences. This report constitutes a part of this work and is the internationally adapted version of the original report in Swedish, published in 2007 (Andersson et al. 2007). The objective of the report is to characterize the genetic variation in Swedish populations of wild plant and animals from the perspective of several themes. These themes were chosen to illustrate general issues currently identified in the international research field of conservation genetics, exemplified with relevant genetic studies of Swedish organisms.
The aim of this report is not to summarize all studies of genetic variation of wild animals and plants performed in Sweden. Studies performed before 1997 have been reviewed in two earlier reports, one about genetic monitoring (Laikre & Ryman 1997) and the other about genetically distinct populations in Sweden (Lönn et al. 1998). These reports are still of immediate importan-ce. Because of the amount of genetic research performed in Sweden and other countries, the present report is not an exhaustive review of all recent progress in the field. Instead it focuses on illustrative examples and on relevant proces-ses that may change the genetic diversity of wild populations in Sweden.
Benefits of genetic variation. The genetic differentiation between
individu-als is the basis of evolution and adaptation. If all individuindividu-als of a species were identical, the species could not adapt genetically to a changing environment e.g. climate change anticipated due to anthropogenic emissions of greenhouse gases. To measure genetic variation and relate it to evolutionary change and ecological function in wild populations is challenging. Recently, some studies have revealed direct effects associated with the genetic variation of a popula-tion. A review summarising the results from several plant-studies shows a
positive correlation between population size, genetic variation and different measures of viability and fertility. Experiments with artificial plant communi-ties having different levels of genetic diversity have also produced evidence. Eelgrass stands consisting of many genotypes were less sensitive to environ-mental change, grew denser, and had larger numbers of small animals associ-ated with them than stands with low levels of genetic diversity.
Human activities may have negative consequences for the genetic diversi-ty, and consequently for the adaptive potential, of wild populations. Regard-less of whether the genetic variation is useful or not in the present-day popu-lations, we cannot anticipate which traits will be essential for survival in a rapidly changing environment.
Loss of genetic diversity in small populations. Decreasing population size
is a problem for many animals and plants that inhabit areas affected by human activities. Small isolated populations are expected to lose genetic diversity through local random processes (genetic drift). Loss of genetic diver-sity may ultimately have effects on the ability to cope with environmental changes (evolutionary potential). Studies show that many populations are so small and isolated that they will be affected by the loss of genetic diversity. Furthermore, populations with low genetic diversity may have reduced fertili-ty and viabilifertili-ty either as a result of inbreeding, or because valuable alleles have been lost. Several studies suggest that a few immigrants are sufficient to eliminate or reduce the negative effects of inbreeding. However, so far it is difficult to draw general conclusions about how the loss of genetic diversity affects the long-term evolutionary potential of populations.
Genetic diversity after gene flow and hybridisation. Although
anthropo-genic habitat fragmentation usually causes negative isolation effects, human activities may also result in increased gene flow between natural populations of animals or plants. A too extensive or too distant gene flow can have nega-tive effects on the recipient populations. Occasionally, human activities have increased the gene flow by creating zones or ”hybrid environments” where genetically dissimilar populations or closely related species can meet and exchange genes. In several species it is known that genes from introduced or domesticated species have spread to Swedish populations, as in the cases of the mountain hare (genes from the brown hare) and two subspecies of the plant lucerne (genes from the cultivated Medicago sativa spp. sativa to the wild relative sickle medick M. sativa spp. falcata). There is only sporadic knowledge about gene flow that occurs when alien populations of trees, birds and fish are released into the wild and come into contact with indigenous populations, or when foreign grass seeds are sown on road verges. Studies of salmon show that gene flow can be harmful by creating hybrids with low levels of viability and fertility (i.e. outbreeding depression). In other cases gene flow has been so intense that the genetic integrity of species is threatened as in the case of the low-density populations of the plant Viola alba on the island of Öland, a species that easily hybridises with other related species. Sometimes populations with the capability to invade natural ecosystems have been created due to human-mediated hybridisation between closely related species. Potentially negative effects of gene flow must also be taken into
con-sideration in conservation biology, e.g. when populations are supplemented with individuals raised in captivity or with individuals from distant popula-tions.
Genetic effects of harvesting. Many Swedish animal and plant species are
exposed to regular harvesting, such as fishing, hunting and forestry. Harves-ting is expected to increase the random loss of genetic variation by decreasing the effective population size. In a study of cod (Gadus morhua), local harves-ting led to increased migration of individuals (and genes) from nearby popu-lations, which also resulted in a change of the large-scale pattern of genetic variation. There are many examples of harvested animal populations that have undergone directional evolution as a result of selective harvesting. In several cases this change has decreased the ability for the population to reco-ver after a period of intense harvesting.
Genetic diversity and change of habitat and climate. Local adaptation
occurs when populations become genetically adapted to different environ-ments. Generally, the ability to adapt is larger the more genetic variation is present in a population. There are many examples of species in Sweden with locally adapted populations in certain environments, e.g. the rough periwink-le (Littorina saxatilis), the common mussel (Mytilus edulis), herring (Clupea
harengus), three-spined stickle-back (Gasterosteus aculeatus), Scots pine
(Pinus sylvestris) and white clover (Trifolium repens). Local adaptation impli-es that individuals from different populations are not interchangeable – locally adapted populations have a conservation value of their own. For this reason, local adaptation is an important issue in e.g. reintroduction and sup-plementation strategies. For each type of adaptation, specific genetic varia-tion is needed. How this variavaria-tion is distributed and exchanged between populations through gene flow is largely unknown.
Genetically distinct populations in Sweden. There are few endemic taxa at
the species level in Sweden, and those that exist have arisen relatively recently through local processes such as hybridisation and polyploidisation. At the same time there are many genetically distinct populations in Sweden. Popula-tions are different due to different origins and colonisation routes or because they are adapted to their local environments. Taxonomic units as species, varieties and forms, together with informal genetic entities such as evolutio-nary significant units and management units reflect genetic differentiation that has arisen within or outside the borders of Sweden. Genetically differen-tiated groups can be difficult to distinguish morphologically (they are cryp-tic), but molecular genetic studies have provided strong evidence for ”hid-den” genetic structure in Swedish taxa. Lönn et al. (1998) called attention to the fact that populations with their main distribution in Sweden are not mar-ginal populations from a genetic perspective. In contrast, species that are mainly distributed in southern areas and are represented by marginal popula-tions in Sweden are less genetically variable in this region. In many cases, Swedish populations are as genetically diverse as the populations in areas that were not covered with ice during the last glaciation. This pattern is also con-firmed by more recent studies. Furthermore, recent investigations also verify that populations from the islands Öland and Gotland, the Baltic Sea with the
surrounding coastal areas, the mountain areas and some traditionally mana-ged landscapes, are genetically distinct. Each population that is lost means loss of genetic variation and consequently loss of adaptive potential. For the purpose of conserving genetic resources, populations or groups of popula-tions are the natural conservation units, because genetic variation occurs both within and between populations.
Genetic monitoring. The need for a genetic monitoring programme in
Sweden was the main conclusion by Laikre & Ryman (1997). We concur that a centrally organised genetic monitoring programme is still needed. Our sug-gestion is largely based on the proposal made in 1997 and is meant as an updated starting point for more detailed discussions of programme design. We propose that the monitoring programme should focus on six different types of taxa (species or groups of populations within species), for example taxa with negative population trends and taxa that are harvested by humans. One of the most important issues for the monitoring programme is to
establish procedures for collecting and storing different types of biological material, e.g. tissue samples, which can be used in genetic investigations. It is very important that the storing of the biological material does not in any way limit which methods that can be used in future investigations. We suggest the establishment of a common database for biological material stored in diffe-rent museums, and we also suggest a formalised system where researchers can report when collected biological material no longer can be stored locally and therefore could be offered to the museums.
Lack of knowledge and suggestion for research areas. In spite of the fact
that many genetic studies have been performed within the focal research field of this report, we have identified several issues with significant knowledge gaps. We would like to see more genetic research regarding the following general questions:
(i) Global warming is likely to result in large changes for Swedish popula-tions and ecosystems. Processes like gene flow and local adaptation will be important for Swedish populations in order to meet these changes. We need more knowledge to address questions such as: What role may genetically distinct populations in Sweden play to enable species to meet large-scale cli-matic and environmental changes? Is there sufficient genetic variation in rele-vant ecological traits to enable species to adapt to rapidly changing environ-ments? Which populations are most valuable in this respect – central popula-tions or those at the periphery of the distribution? Is there a risk that geneti-cally distinct populations will disappear in those habitats that supposedly will be most affected by global warming? Which genetic methods are most rele-vant for assessing evolutionary potential? How will genetic variation of key species in important Swedish ecosystems affect the function, species composi-tion and stability of these ecosystems?
(ii) It is important to understand how the human-mediated gene flow affects the gene pool of the genetically distinct populations present in Sweden. What genetic effect does the release of alien populations that takes place in e.g. the forestry and fisheries have? How will species of natural grass-lands be affected by the gene flow from foreign provenances sown on road
verges? When is it appropriate to supplement small or inbred populations, or populations that are poorly adapted to a changing environment? When will such measures be harmful? From which populations should the individuals used in the supplementation be taken?
(iii) There are several scientific questions that are in need of answers when developing a genetic monitoring programme. One of the most central ques-tions is: How can we differentiate natural changes in genetic diversity from the un-natural, which might constitute a threat to the genetic diversity?
What should the national action plan include? The national action plan
for conserving genetic diversity in wild plants and animals in Sweden should first and foremost include a genetic monitoring programme and also prioriti-se the suggested reprioriti-search areas. Furthermore we suggest that the action plan proposes guidelines for how to deal with genetic problems in monitored populations and in the supplementation of wild populations, an action alrea-dy suggested in several of the recovery and action plans for red-listed species in Sweden. The national action plan should also aim to develop guidelines and effective systems for the registration of the release of alien populations that take place e.g. in forestry, fish management and in road management.
1. Background
Conservation and sustainable use of biological diversity is presently a politi-cal objective for many countries in the world. The International Union for the Conservation of Nature and Natural Resources (IUCN) identifies three levels of biological diversity that are equally important to conserve: ecosystem, spe-cies and genetic diversity (McNeely et al. 1990). The UN Convention of Bio-logical Diversity (CBD) makes a similar classification. Until now, the conser-vation of genetic resources has attracted relatively little attention in practical nature conservation. One common assumption is that the conservation of ecosystems and habitats also conserves species, and that the conservation of species also conserves genetic diversity. However, there is a growing realiza-tion that the conservarealiza-tion of species does not necessarily conserve the genetic diversity within them. Within the CBD there is an agreement to work from the general perspective of the ecosystem level. They suggest that the biological diversity should be viewed from the landscape perspective, which also inclu-des socio-economic factors, but emphasize that this does not contradict actions directed to conserve biological diversity on either the species or the genetic levels.
Genetic differences between individuals are the basis of evolution and adaptation. If all individuals of a species were identical the evolution would be disabled, all individuals would for example be equally sensitive to a certain disease or would react to an environmental change in the same way. As a con-sequence the species could not adapt genetically to a changing environment e.g. climate change due to anthropogenic emissions of greenhouse gases. In spite of the limited attention the conservation of genetic diversity has received within practical conservation biology, knowledge of the importance of gene-tic variation for the survival of species is far from new. Already Darwin (1896) understood the importance of genetic variation when he pointed out that the reason why deer in British parks were in bad condition was because they were kept in small isolated populations (see also Allendorf & Luikart 2007). However, it was not until after 70 years the actual debate about the conservation of genetic resources started with a paper in the journal “Gene-tics” (1974) by Otto Frankel, an agricultural plant geneticist. Frankle (1974) argued that because we cannot anticipate what the future world will look like, we cannot predict which traits the organisms will need to survive there. For this reason it is vital that the level of genetic variation within species is conserved so evolution may continue (Frankel 1974; see also Allendorf & Luikart 2007).
In a society with limited economical resources, conservation actions often are weighted against each other and against other interest of society. Which species should we conserve? Should the economic resources be used to con-serve natural habitats instead of species? In recent years several scientists have argued that the conservation of biological diversity should focus on preven-ting the disappearance of genetically distinct populations rather than to solely
prevent the extinction of species (Hughes et al. 1997; Hobbs & Mooney 1998). To conserve genetically distinct populations could be a better way of conserving the evolutionarily potential of species. This will also reduce the risk that species go extinct, even in a longer time perspective (Allendorf & Luikart 2007).
A population focus in conservation biology suggests that Sweden together with the other Nordic countries share a common responsibility to conserve the populations in this area. Many species also present in continental Europe are in Sweden represented by so-called marginal populations – populations at the edge of their distributions. These populations are often genetically distinct and are therefore essential to protect if we aim to conserve the evolutionary potential of a particular species.
The importance of conserving genetic diversity is emphasised in one of the Swedish environmental objectives “A Rich Diversity of Plant and Animal Life”, which was adopted by the Swedish parliament in November 2005. In 2006, the Swedish Environmental Protection Agency received a government commission to develop a national action plan for the conservation of genetic diversity in wild plants and animals, in consultation with the Swedish Board of Agriculture, the Swedish Forest Agency, the Swedish Board of Fisheries and the Swedish University of Agricultural Sciences. This report constitutes a part of this work and is the internationally adapted version of the original report in Swedish, published in 2007 (Andersson et al. 2007).
1.1 The aim and focus of the report
The objective of this report is to characterize the genetic variation in Swedish populations of wild plant and animals from the perspective of several themes. These themes were chosen to illustrate general issues currently identified in the international research field of conservation genetics, exemplified with relevant genetic studies of Swedish organisms. In this section we describe the background and aim of this report in more detail.
Even if the genetic aspect of practical conservation biology has not always been prioritised, the question has attracted some attention in the Swedish Environmental Protection Agency since the late 1990s. At that time two reports were published that reviewed the genetic variation of natural plants and animals in Sweden from two, slightly different, perspectives.
The first report discusses the monitoring of biodiversity at the gene level (Laikre & Ryman 1997), and suggests an action plan and a research pro-gramme to ensure that the genetic diversity of Swedish organisms is conser-ved. The authors emphasise the need for long-term studies of natural fluctua-tions of genetic variation to be able to differentiate between natural and un-natural changes of genetic diversity. The report also lists practically all
molecular genetic studies of natural plants and animals in Sweden, performed until November 1996. The list was created in part through literature searches in databases of scientific studies and in part by a questionnaire sent directly to various research institutes, and identifies 316 studies. Although many studies
of genetic variation of Swedish organisms exist, Laikre & Ryman (1997) emphasise that these studies are not necessarily useful when for example assessing the conservation status of a species from a genetic perspective.
The second report discusses the genetic distinctness of Swedish popula-tions of natural plants and animals in relation to populapopula-tions in other Euro-pean areas (Lönn et al. 1998). This report reviews mainly molecular, but also some quantitative genetic studies, and presents many studies illustrating that Swedish populations are genetically distinct. Lönn et al. (1998) describe seve-ral geographic areas in Sweden that contain many genetically distinct (mostly plant) populations: the mountain areas, the Baltic Sea with the surrounding coastal areas, the Baltic sea islands Öland and Gotland and some traditio-nally managed landscapes. Because taxonomic and genetic variation does not always coincide, the authors stress the importance of performing genetic stu-dies in order to discover genetic distinctness.
These two reports refer to a large number of genetic studies of natural plant and animal populations in Sweden, and together they provide an exhau-stive review of the genetic studies published before 1997. Both reports are still of immediate importance, because much of the information in them has not significantly changed over time.
Since 1997 a large number of genetic studies of wild organisms in Sweden has been published. Previous to writing the present report, a literature search in three different databases of scientific studies (Biological Abstacts, ISI Web of Knowledge, CSA Illumina) was performed for the years 1996 to November 2006, using the same key words as in Laikre & Ryman (1997). The search resulted in the identification of 844 studies of genetic variation of Swedish organisms. However, the aim of the present report is not to summarize all stu-dies of genetic variation of wild animals and plants performed in Sweden sin-ce 1997. For some organism groups, or specific questions, reviews of this kind are already published. For example a report of genetic research of com-mercially exploited fish species in the Nordic countries was published recent-ly by the Nordic Council of Ministers (Olsson et al. 2007). Other reports discuss the problem of release of genetically alien populations in Sweden (Laikre & Palmé 2005; Laikre et al. 2006) and research concerning ecological effects of genetically modified organisms (Palm & Ryman 2006). The present report is instead a complement to these more detailed reports, and focus on the processes that modify the genetic variation of natural populations descri-bed from a Swedish perspective. Genetic variation of Swedish natural popula-tions of plants and animals is described from the perspective of several the-mes, chosen to demonstrate what problems and general threats to their gene-tic diversity that natural populations presently are facing. Each chapter corre-sponds to a theme, and opens with a short summary followed by some information about the theoretical background and results from international research, after which examples are provided of Swedish research relevant to the issue at hand. Some of the questions addressed in this report, and discus-sed from the perspective of Swedish genetic studies, are:
• Why do species need genetic variation?
• What are the negative consequences of gene flow and hybridisation? • Can harvesting of natural populations, for example by fishing and
hun-ting, have genetic consequences
• How will the ongoing climate changes affect natural populations in Swe-den?
• What should the national action plan for the conservation of genetic diversity in wild plants and animals in Sweden include?
1.2 Basic theory of population genetics
The present report is the English adaptation of a report previously published in Swedish by the Swedish Environmental Protection Agency (Andersson et al. 2007). In addition to the objective of describing problems and general threats to the genetic diversity of Swedish plants and animals, the original report also had a second equally important aim – to give a presentation (in Swedish) of relevant parts of the basic theory of population genetics for peo-ple who are less familiar with the subject, or those who wished to renew their theoretical knowledge (Figure 1). The Swedish version of this report also included an appendix where the most common molecular genetic methods were described, summarising the advantages and drawbacks of current molecular methods. The reason for not including these parts in the internatio-nally adapted version of the report is that there are several excellent books in English, describing basic population genetic theory, evolutionary processes and current molecular methods. If you are interested in this subject or want to renew your theoretical background we recommend you to read for examp-le “Conservation and the Genetic of Populations” (Alexamp-lendorf & Luikart 2007) or “Introduction to Conservation Genetics” (Frankham et al. 2005), or some other textbook of the subject.
Chromosome Copy of the chromosome Cell Cell nucleus Mitochondria Nuclear DNA Mitochondrial DNA Supportive proteins Base pairs
Figure 1. In an animal cell DNA is found in the nucleus of the cell and in the mitochondria (the “power plants” of all eukaryotic cells). In a plant cell DNA can also be found in the chloroplasts. Nuclear DNA is accumulated in chromososomes, which consist of a extremely long DNA molecules wrapped around different types of proteins. During most of the cell cycle, the chromosomes are generally loosly packed, and are streched out in long threadlike formations in the cell nucleus. Before cell division, the DNA is replicated to create identical copies of all the chromososmes. After the replication all chromosomes condense, i.e. they are tightly packed around the supportive prote-ins. At this time the chromosome can be stained and studied using a microscope. Becaue the origi-nal chromosome and its identical copy are attached to each other, the condensed chromosome often has a characteristic X-shape.
Mitochondrial DNA (mtDNA) is a circular molecule that exists in several copies inside each mitochondrion. The organisation of mtDNA is not as well known as nuclear DNA. A study of human mitochondria demonstrated that one or probably several mtDNA molecules are loosely organised together with (unknown) proteins into loosely packed structures (Garrido et al. 2003).
Illustration: A-C Lundqvist. The illustration is rewritten and modified from an illustration on the homepage, www.genome.gov (The National Human Genome Research Institute, NHGRI, Educatio-nal resources, Talking glossary)
2. Benefits of genetic diversity
2.1 Why should we conserve genetic variation?
Genetic differentiation between individuals is the basis for the evolutionary change of species, populations and lineages. In which way a population or species will change is determined by natural selection – individuals with cer-tain traits will survive to a greater degree and/or will produce more offspring. This evolution will result in a slightly altered genetic composition of the spe-cies or population. A spespe-cies or population may also evolve genetically due to a random process, genetic drift. Random genetic drift changes the frequency of alleles (alternative forms of a gene) between generations; some alleles will decrease in frequency while other will become more common, only depending on random effects during the formation of gametes (e.g. egg or sperm cells). However, random genetic drift generally is a weak force and evolution through this process will not modify a population to any great extent. This will require a directed force such as natural selection.
Evolutionary change requires modification of genes or gene combina-tions. The functions of existing alleles are changed through mutation or recombination. Favourable mutations may enable the organisms to better uti-lise the existing environment – predators may develop the ability for more rapid attacks, whereas prey might develop a more acute sense for detecting the predators.
If the environment changes, for example if a predator invades a new habi-tat or if the climate suddenly changes, yet another situation can arise. Alleles that previously were neutral or had a small effect on the reproductive success of the individuals suddenly can become important. Alleles controlling frost tolerance will be of no importance in an environment that never experiences temperatures below zero, but can rapidly become important if the climate
Summary:
The genetic differentiation between individuals is the basis of evolu-tion and adaptaevolu-tion. To measure genetic variaevolu-tion and relate it to evo-lutionary change and ecological function in wild populations is chal-lenging. Recently, some studies have revealed direct effects associated with the genetic variation of a population. A review summarising the results from several plant-studies shows a positive correlation between population size, genetic variation and different measures of viability and fertility. Experiments with artificial plant communities with diffe-rent levels of genetic diversity have also produced evidence. Eelgrass stands consisting of many genotypes were less sensitive to environ-mental change, grew denser, and had larger numbers of small animals associated with them than stands with low levels of genetic diversity.
suddenly turns colder. Nevertheless, obtaining useful genetic variation through mutation is a very slow process. Consequently, the probability that a population will survive a forthcoming environmental change will increase the more alleles that are present in the population because some of the alleles might be useful just by chance.
It is impossible to anticipate which alleles will be needed for a species to survive in the future. Obviously, the probability that a population or species has the required allele increases with the amount of genetic variation that is present in the population. For a specific population to survive, at least one individual (but preferably many individuals) must possess the allele. In the same way, for a species the allele has to be present in at least one of the popu-lations. If only one or a few populations possess the necessary allele, other populations can receive it through gene flow or because a population disap-pears and is replaced by individuals from another more viable population possessing the allele. However, the more populations that have the required allele the better – the distance between populations can some times be consi-derable and for many organisms dispersal is limited to small distances (10-1000 meters; see Edenhamn et al. 1999). It is essential to possess many alleles (high allelic richness), but it is also important that the different alleles are rea-sonably common (high levels of allele diversity), because that decreases the probability that alleles are lost through random processes.
To measure genetic variation and relate it to evolutionary change and eco-logical function in wild populations is challenging. First, the genotypes of the individuals in the populations under study must be known. Second, an envi-ronmental change affecting the success of the studied alleles must occur (because it is not possible to study all alleles at all loci in a population). After that the relative success of the different populations or lineages can be evalua-ted based on their genetic variation previous to the environmental change.
Genetic variation present in a population today may not be entirely bene-ficial in the current environment. A few alleles may be directly harmful for the individuals carrying the alleles under the present circumstances. Several examples of this have been recorded in natural populations, when organisms have adapted rapidly to "unexpected" environmental changes, as for example deposits of toxic heavy metals or other pollutants. Frankham & Kingsolver (2004) review a number of scientific studies demonstrating this phenomenon in natural populations. One classical example is colour adaptation in the pep-pered moth (Biston betularia), a species that resides on the trunks of light-coloured trees in Britain. During the industrial revolution pollution discolou-red the trees, and the dark colour of the trunks was reflected in an increased proportion dark-coloured moths. The dark, melanistic, variants of the moth were disadvantaged when the tree trunks were pale, but natural selection made the alleles causing the melanistic form much more common because the environment had changed. Years later, when the pollution was less pronoun-ced the light-coloured moths became more common once more, at the expen-se of the darker variant.
Several examples from other groups of organisms also show this pheno-menon. In many cases, plants that grow in localities where mining residues
are deposited will adapt to the high levels of heavy metals and become resi-stant to them. Also, Swedish populations of the moor frog (Rana arvalis) have rapidly adapted to the acidified environment caused by atmospheric pol-lution (Räsänen et al. 2003). However, this is not true for all species and populations, because some of them lack the alleles that enable adaptation to the novel environment (Davies 1993).
In general, natural populations show high levels of genetic variation whi-ch can be expressed if the selection pressures whi-changes. Plant and animal bree-ding are based on this genetic variation. By artificially altering the selection pressure (this corresponds to an environmental change) populations rapidly gain traits because some of the alleles are “favoured”. One experiment using fruit flies (Drosophila melanogaster) showed that populations kept in an unpredictable environment were more successful when they were exposed to an environmental change compared with populations that had been kept in a stable environment (Reed et. al. 2003). The authors suggest that populations kept in a variable environment have the possibility to maintain higher levels of genetic diversity and will consequently have the capacity to face unknown environmental changes.
2.2 The positive effects of genetic variation
at the population, community and ecosystem
levels
Benefits of genetic variation can be shown for example by the ability of diffe-rent populations to adapt to their local environment; this will be discussed in a different section (Chapter 6). The main focus of the present section is studi-es examining the level of genetic variability within specistudi-es or populations (i.e. genetic diversity of populations).
If we assume that the levels of genetic variation shown by molecular mar-kers reflect the variation of the entire genome, the benefits of genetic varia-tion can be studied indirectly using molecular markers. A meta-analysis (a summarising analysis of the results of several scientific studies) showed that population size, genetic variability and different fitness related traits (fertility and viability) all were positively correlated with each other. Thus, in this stu-dy the benefit of genetic variation was demonstrated by its positive associa-tion with fitness. The authors stress that because populaassocia-tion size and genetic variability also are positively correlated, habitat fragmentation may pose a risk to the genetic diversity of populations, as a consequence of the reduced populations size and limited gene flow between the subpopulations (Leimu et al. 2006).
Recently, a newly emerged research field termed “community genetics” has received some scientific attention. Community genetics is the study of how the genetic variability of one species in a community or ecosystem, affects other species or ecosystem functions. To investigate if the genetic vari-ation of the eelgrass (Zostera marina), a dominant plant in shallow waters,
would have an effect on the function of the entire ecosystem, Reusch et al. (2005) performed a manipulative field experiment in the Baltic Sea. Eelgrass was planted in three different groups either consisting of one, three or six eel-grass genotypes. During the experiment, a natural period of extremely high (for the eelgrass close to lethal) water temperatures occurred. Thus, the expe-riment also reflected the resistance to environmental stress of the manipulated eelgrass communities. The results showed that eelgrass in communities consi-sting of several different genotypes grew at a higher density and produced more biomass than those with lower diversity. Genetically variable communi-ties performed better, either because the different genotypes facilitated their mutual growth or because the different genotypes utilised the environment in slightly different ways. Genetically more variable eelgrass communities were also associated with a greater number of small animals, which are a valuable food source for larger animals in the ecologically important eelgrass eco-systems. This study shows that genetic variation of a species can have far-reaching consequences both for other species and for the entire ecosystem.
For individual organisms, the importance of genetic variation can be demonstrated by the fact that different genotypes do not have identical fitness in different environments – this is the basis of natural selection. At the popu-lation level, the importance of genetic variation can be shown by demonstra-ting that the success of a population depends on the genetic variability within the population. In a study of the fastigiate gypsophila (Gypsophila fastigiata), a rare plant in the genus baby’s breath on the Baltic island of Öland, the gene-tic variation was compared between different subpopulations from different areas in the regional population (Lönn & Prentice 2002). After correcting sta-tistically for geographic position of the subpopulations, it was shown that genetically more variable subpopulations displayed a higher frequency of young individuals, but did not show an elevated number of dead individuals, which indicates that these populations are expanding at the expense of other subpopulations. This can be explained by the fact that in the genetically more variable populations, each individual displays higher levels of genetic varia-tion (are heterozygous at a large number of loci), which in turn enables each individual to survive in areas showing a wide span of environmental varia-tion. Another explanation could be that each genetically variable population can generate many different genotypes, which together can utilise the envi-ronment throughout the entire distribution area in a more effective way.
In a manipulative field experiment performed on an English grassland, genetic variation was found to be an important issue for plant community diversity (Booth & Grime 2003). In this experiment, the ten most common species were used to construct artificial plant communities with three levels of genetic diversity. Each species was represented either by a number of geneti-cally identical individuals (1 genotype) or by a number of individuals of 4 dif-ferent genotypes, or by individuals with unique genotypes (16 difdif-ferent geno-types). The results demonstrated that the genetically variable artificial plant communities were able to recreate plant communities resembling the natural grassland. In contrast, the artificial plant communities where each species consisted of genetically identical individuals were very different from each
other, and from the original grassland, even after a long time period (Figure 2). These results indicate that different genotypes may have a different func-tion in the interacfunc-tion between species, and the authors suggest that a mature plant society not only has a certain species composition, but also a specific composition of genotypes.
Furthermore, the species diversity declined much more slowly in the gene-tically variable artificial plant communities, which might be due to decreased competition between the different species or an elevated resistance to disease.
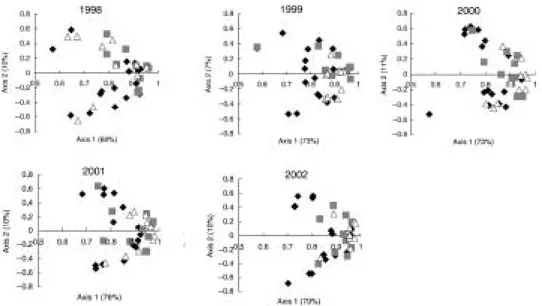
Figure 2. Ordination of the species composition over 5 years in 36 artificial plant communities. Each diagram shows a principal component analysis. An ordination arranges object according to likeness. The closer the dots are to each other the more the species composition in the different communities resembles each other. White triangles = communities where each species is represen-ted by 16 genotypes; grey squares = 4 genotypes; black diamonds = 1 genotype. From: Booth, R. E. & Grime J. P. (2003): Effects of genetic impoverishment on plant community diversity. Journal of Ecology 91: 721-730. Copyright and permission to publish: Wiley-Blackwell, Oxford.
Prentice et al. (2006) observed a related process at the species level, study-ing the quakstudy-ing-grass (Briza media) on the Baltic island of Öland (Figure 3). The study area is a grassland with a continuity of up to 300 years. Popula-tions of quaking-grass were found to be genetically more similar (measured with allozymes) the longer the grassland continuity at the specific locality was. The authors suggest that the environment and the species composition become more homogenous in areas with long grassland continuity, and that specific genotypes might be better adapted to that plant community (i.e. directional selection may favour optimal genotypes over time).
One of the main conclusions both from Booth & Grime (2003) and Pren-tice et al. (2006) is that it seems to matter which genotypes represent a species in a community. Therefore, it is important to consider the genotypes of indivi-duals in conservation actions such as habitat restorations, or when predicting changes in the distribution of species or in studies of community and eco-system processes.
Figure 3. Quaking-grass (Briza media) is a characteristic grass species in pastures in Sweden. (Photo: Myra bildbyrå)
3. Loss of genetic diversity in
small populations
Most natural populations have at least moderate levels of genetic diversity – a basic condition to enable species and populations to adapt to the present and future changes of the environment. Since the 1980s conservation biology often has focused on the loss of genetic variation that is expected in small iso-lated populations or populations experiencing "bottlenecks" – time periods when the size of the population is very small. Loss of genetic diversity has long been considered as a problem mostly concerning rare plants and ani-mals, but this loss can also – and sometimes to a greater degree – have an effect on common species with large populations, especially species affected by the ongoing fragmentation of the landscape (Ellstrand & Elam 1993; Frankham 1995a; Young et al. 1996). In addition, low levels of genetic diver-sity can often be observed in populations inhabiting areas that were recoloni-sed only after the latest ice age (Hewitt 1996).
Populations can show low levels of genetic diversity or be genetically impoverished in different ways: (1) many alleles may have become rare or are absent from the gene pool because of genetic drift; (2) the proportion of homozygotes may have increased at the expense of the heterozygotes due to
Summary:
Decreasing population size is a problem for many animals and plants that inhabit areas affected by human activities. In this chapter we discuss the effect of loss of genetic diversity in small isolated popula-tions due to local random processes (genetic drift). In the beginning of the chapter we consider the loss of genetic diversity at the individual level, which can be studied using different molecular methods. Then we discuss how loss of genetic diversity in a population may ultimately have effects on its viability, fertility and ability to cope with environ-mental changes (evolutionary potential). Each section starts with a short theroetical backgroud followed by examples from empirical stu-dies, focusing on studies of Swedish plant and animals.
Studies show that many populations are so small and isolated that they will be affected by the loss of genetic diversity. Furthermore, populations with low genetic diversity may have reduced fertility and viability either as a result of inbreeding, or because valuable alleles have been lost. Several studies suggest that a few immigrants are suffi-cient to eliminate or reduce the negative effects of inbreeding. Howe-ver, so far it is difficult to draw general conclusions about how the loss of genetic diversity affects the long-term evolutionary potential of populations.
inbreeding in the population; (3) the gene pool may be fixed for alleles that decrease the viability and fertility of the individuals or the population.
3.1 Loss of genetic variation on the
individual level
Many populations are so small that a random process, genetic drift, has a sig-nificant impact on the number of alleles that are passed on to the next genera-tion. Genetic drift has two major consequences: the frequencies of the diffe-rent alleles will fluctuate between generations, and the population will loose genetic variation. Initially, the rare alleles will be lost, and with time the ave-rage heterozygosity (the allele diversity) will decrease. Genetic diversity will decrease within the populations while the genetic differences between the populations may increase or decrease, depending on whether the random genetic drift acts in the same or opposite directions in different populations. The genetic diversity (both the loss of rare alleles and the average heterozygo-sity) within the populations will decrease at a rate inversely proportional to the effective population size (Ne). The average heterozygosity within the population is expected to decrease at rate of 1/2Ne per generation (Box 1). This loss will continue until the gene pool is fixed for one allele or until an equilibrium arises between the loss of genetic variation and creation of gene-tic variation through mutations (Wright 1931; Kimura 1968).
The effect of genetic drift is easiest to illustrate with an example of a population that has gone through a short period of small population size. If a large population decreased to an effective size of Ne = 20 individuals, and maintained this size for five generations, the population is expected to have 88 % of the allele diversity (expected heterozygosity) present before the size reduction. If the population instead decreased to an effective size of one indi-vidual (Ne = 1), and that indiindi-vidual establishes a new population through self-fertilization, this population is expected to retain only half of the original allele diversity of the first generation of the bottleneck. An extreme bottle-neck, like the latter example, will also drastically reduce the number of alle-les, because one diploid individual can only pass on at most two different alleles at each locus to the next generation. In general, it is the rare alleles that will be lost in bottlenecks (Nei et al. 1975).
Only a small amount of gene flow (gametes, pollen, migrating individuals) is needed to counteract the loss of genetic diversity in a population. In a group of populations exchanging genes between each other from time to time an equilibrium is expected to arise, where the effects of genetic drift are balanced by mutations and gene flow. This equilibrium is expected to arise quickly for the various estimates of genetic differentiation between popula-tions (GST, FST), but more slowly for estimates of genetic diversity within
populations (allele diversity). As a consequence, estimates of allele diversity of populations in a recently fragmented landscape will indicate their larger “historical” population sizes and the estimates of allele diversity will decrease
further, before an equilibrium between gene flow and mutations will be reached (Wright 1931; Varvio et al. 1986).
Box 1: Loss of genetic variation
In all finite populations, genetic drift is expected to decrease the genetic variation in a population every generation. According to population genetic theory it is possible to show that a finite population will loose a certain proportion of the average heterozygosity (allele diversity), becau-se the proportion of homozygotes in the population will increabecau-se due to inbreeding. The increased inbreeding, or the increase of the inbreeding coefficient (F), can be expressed as:
where Ne is the effective size of the population.
Inbreeding increases the proportion of homozygotes, because mating between closely related individuals increases the frequency of homozy-gous loci in each offspring compared with mating between non-related individuals. The homozygotes (or homozygous loci) generated by inbreeding will consequently have two identical alleles that can be tra-ced back to the same individual – the alleles are identical by descent. If we consider the total gene pool of a population of N diploid individuals, the probability to "pick" the same allele twice is 1/2N for any allele in the gene pool. This can be explained as a process in two steps: first one specific allele is randomly picked from the gene pool and in the second step the exact same allele is picked a second time. Because the frequency of this specific allele is 1/2N in the gene pool (the allele only exists in one single copy), the probability to pick this specific allele in the second step is 1/2N.
If the frequency of homozygotes increases with a factor 1/2Ne the expected frequency of heterozygotes (allele diversity) will at the same time decrease with the same factor. A population of 10 individuals will each generation loose 1/(2x10) = 0.05 = 5 % of its heterozygosity.
Conservation geneticists have tried to define population sizes that are large enough to retain sufficient genetic variation (1) to avoid loss of reproductive fitness or (2) to enable the population to evolve in response to environmental changes. Soulé (1980) and Franklin (1980) argued that an effective population size of Ne = 50 (1% of genetic variation is lost each generation) is sufficient to retain reproductive fitness, and
Ne = 500 is sufficient to retain evolutionary potential because 0.1% of
genetic variation is lost each generation, and this loss will be
compensated by new genetic variation created by mutation. However, this approach is under much debate, and there is no consensus among conservation geneticists which population sizes that are “safe” from a genetic perspective. e
N
F
2
1
=
Δ
3.1.1 Studies of natural populations
A meta-analysis including published studies of 102 species (of which most were animals), showed that estimations of the effective population size (Ne) on average is about 10 % of the census size (N), and that many populations are so small that they might be affected by genetic drift and/or bottlenecks (Frankham 1995b). In a meta-analysis of 46 enzyme and DNA-studies of 41 plant species, Leimu et al. (2006) demonstrated that small populations gene-rally have lower frequencies of variable genes, fewer alleles per locus and lower levels of allele diversity compared with larger populations. This explains why rare or threatened species – which often occur in small isolated populations – frequently have less genetic variation than common species, both in relation to the “average” population and the entire species (Cole 2003; Spielman et al. 2004). Molecular markers also confirm that loss of genetic variation leads to an increased random differentiation between popu-lations (Cole 2003).
The hazel (Corylus avellana) demonstrates apparent signs of genetic drift, despite the fact that pollen from this bush is wind dispersed over large distan-ces (Figure 4). Populations in Sweden lack many of the chloroplast-DNA haplotypes that can be observed in southern and central parts of Europe, as a consequence of bottlenecks occurring during the early and rapid recolonisa-tion of Scandinavia after the most recent ice-age (Palmé & Vendramin 2002). There are also results indicating random effects at a later date occurring locally: small isolated populations of hazel growing on south facing hillsides in north Sweden have fewer variable genes, fewer alleles per locus and lower levels of allele diversity (heterozygosity) than the larger continuous popula-tions in southern parts of Sweden (Persson et al. 2004).
Figure 4. Hazel (Corylus avellana) has been studied genetically to investigate the effects of isola-tion and colonizaisola-tion history. To a large extent, Swedish populaisola-tions of hazel lack the genetic varia-tion of chloroplast DNA that can be observed in southern and central parts of Europe. The decrea-sed level of variation is likely an effect of bottlenecks during the rapid recolonisation of Scandinavia after the most recent ice age. A more recent loss of genetic variation in the small isolated popula-tions of hazel in northern Sweden has also been observed. (Photo: Myra bildbyrå)
The loss of genetic variation observed in populations of the flower prolife-rous pink (Petrorhagia prolifera) in southern Sweden is much more pronoun-ced that the moderate loss of genetic variation observed in hazel. The prolife-rous pink is an annual self-reproducing plant that demonstrates a mosaic pat-tern of genetic variation geographically – separate populations have been fix-ed for different allozyme alleles. Almost all genetic variation (>90 %) is found among populations (Lönn & Prentice 1990), whereas the corresponding esti-mate in hazel is 8 % (Persson et al. 2004). The pattern of genetic variation is probably created by bottleneck effects as a consequence of the reproductive system, which enables the proliferous pink to establish a novel population only from a few individuals. Furthermore, there are only slight possibilities that gene flow between populations will compensate for the loss of genetic diversity and counteract the population divergence, because the seeds have no dispersal adaptations and most pollen grains fail to leave the small flowers.
Some species seems to be very resistant to the loss of genetic diversity. One of these, the spring pea (Lathyrus vernus) is a boreal, insect-pollinated plant, lacking specialised adaptations for seed dispersal. The genetic variation at several loci show that marginal populations in north Sweden display the same levels of allele diversity as populations in Central Europe, which are closer to the supposed glacial refuge. In addition, the estimates of allele diversity are similar both in “small” (<150 ind.) and “large” (>500 ind.) populations. The high levels of genetic diversity in small marginal populations of the spring pea can be contrasted with the loss of genetic variation that is observed in popula-tions of hazel growing in similar geographic areas, a phenomenon that might be explained by a slower recolonisation (lack of bottlenecks) in the spring pea (Schiemann et al. 2000).
In recent times, studies of the loss of genetic variation have largely been focused on natural populations of animal and plant species that previously had large continuous distributions but where the present populations are small and isolated due to the ongoing fragmentation of natural habitats (Jac-quemyn et al. 2004; Keyghobadi et al. 2005; Honnay et al. 2006; Prentice et al. 2006). Because the fragmentation often is recent, there are often maps or aerial photos available which can be used to show how the different frag-ments were connected and how large the fragfrag-ments were originally. This information can be combined with data from molecular genetic markers, to demonstrate how fast the loss of genetic variation affects natural populations. Several of the species suffering from fragmentation show apparent signs of loss of genetic variation, whereas the genetic variation in other species are still typical of the larger population sizes that existed in the previous continu-ous landscape (Keyghobadi et al. 2005). In contrast, other studies indicate that the fragmentation of habitat occasionally may counteract the loss of genetic variation because gene flow from very distant populations may incre-ase, introducing novel genetic variation in the gene pools (Young et al. 1996; White et al. 2002).
Recently bottlenecked populations often show low levels of genetic varia-tion in molecular markers. This can be seen for example in animals and plants that have recovered from near extinction e.g. the northern elephant seal
(Mirounga angustirostris; Hoelzel et al. 1993), Mauritius kestrel (Falco
puncta-tus; Groombridge et al. 2000) and the Swedish beaver and wolf populations
(Castor fiber, Canis lupus; Ellegren et al. 1993, 1996). In several cases, (e.g. Mauritius kestrel), the level of genetic variation are in agreement with what can be expected from the bottleneck period. In other species, e.g. the northern elep-hant seal, the level of genetic variation is too low to be explained only by the bottleneck. In this particular case, the extremely low levels of genetic variation can be explained by the fact that predators (as a group) often have low levels of genetic variation in molecular markers (Amos & Balmford 2001). This examp-le demonstrates the importance of comparing results to reexamp-levant reference mate-rial, e.g. large populations of the same species (Ellegren 1993, Leimu et al. 2006), or a common species within the same genus (Cole 2003; Spielman et al. 2004), when evaluating the effects of bottlenecks and variation losses.
Figure 5. The current population of wolves (Canis lupus) in Scandinavia was established from a limited number of individuals arriving in the 1980s and 1990s. The population has since expan-ded, but shows apparent signs of both inbreeding effects and low levels of genetic variation. (Pho-to: Myra bildbyrå)
Some bottlenecks are followed by a rapid recovery of genetic variation, most likely caused by a small number of immigrants arriving after the bottle-neck (Keller et al. 2001). An illustrative example of this phenomenon is the Scandinavian population of wolves (Figure 5). This species had been absent from Sweden for a couple of decades, but in 1983 a pack of wolves was obser-ved in southern Scandinavia, more than 900 km from the neighbouring popula-tions in Finland and Russia. The pack consisted of about 10 animals for a num-ber of years, and evidence of loss of genetic variation and inbreeding could be seen in molecular markers (Ellegren et al. 1996). Suddenly, the population expanded and started to disperse to other geographic regions in the 1990s. Stu-dies using microsatellites showed that the expansion begun with an increase of the genetic variation (increased levels of heterozygosity, several new alleles). The increase of the genetic variation was most likely caused by the arrival of a single male to the Scandinavian population (Vilá et al. 2002). Furthermore,
recent microsatellite data show that it is more common that individuals with high levels of genetic variation breed, which could reduce the negative effects of inbreeding among the wolves in Scandinavia (Bensch et al. 2006).
3.2 Effects on fertility and viability
Random processes such as genetic drift affect all parts of the genome uni-formly, not only neutral loci often used in studies of genetic variation, but also the genes affecting the fertility and viability of individuals (fitness). Con-sequently, loss of variation in molecular markers could indicate that genetic variation in ecologically important traits also has been lost. In this context it is important to keep in mind that “essential” alleles are expected to be pre-sent at high frequencies because they are favoured by selection, and thus they will most likely be present even in small gene pools (Nei et al. 1975). Advan-tageous alleles that are only needed infrequently (e.g. during a disease outbre-ak), will be lost at a much higher rate, because no selective force operates to keep them in the population between these events (Amos & Balmford 2001).
The risk for “serious” allele losses is also large in the so-called S-locus, genes that control self-incompatibility in plants and fungi (Box 2). In general, this system favours rare S-alleles; these will seldom encounter a copy of them-selves and as a result they will have a high probability of being passed on to the next generation.
A population in equilibrium will consequently have a large number of S-alleles present at low frequencies. Therefore, the gene pool of a population at equilibrium will be at great risk to loose alleles if the number of individuals suddenly declines, because random genetic drift is greater in smaller popula-tions. Because of the random loss of S-alleles, many individuals will have identical S-allele genotypes which mean that fewer individuals will be able to mate with each other, thus affecting the reproductive success of the entire population. Computer simulations demonstrate that populations with less than 25 individuals will not be able to keep sufficient S-alleles to maintain a high level of reproductive success (Byers & Meagher 1992).
Box 2: Self-incompatibility locus (S-locus)
The self-incompatibility locus in some plants and fungi is a locus under frequency-dependent selection. The function of this locus is to prevent self-fertilization and breeding between closely related individuals. Gene-tic variation in this locus is a necessity for breeding, because pollen (or spores) with a certain genotype at the S-locus will only be able to fertili-ze individuals that possess at least one different allele at the same locus. A novel S-allele, introduced into the population by migration or muta-tion, will be at a selective advantage because the individual that possess this allele will be able to fertilize all individuals in the population except itself. As a result, the frequency of the novel allele will increase until it reaches the same level as the other S-alleles.
On top of loosing genetic variation as a consequence of genetic drift, small isolated populations are also expected to become inbred because each generation the individuals are continuously getting more related to each other. The inbreeding is entirely an effect of the reduced population size, and will increase with 1/2Ne each generation, i.e. at the same rate as the decrease of heterozygosity and additive quantitative genetic variation (Box 1). Inbree-ding will increase the homozygosity and the proportion of alleles that are identical by descent. Eventually, the proportion of homozygotes will be so large that the population might suffer from inbreeding depression, similar to what occurs after self-fertilisation or breeding between siblings (Charles-worth & Charles(Charles-worth 1987).
With increased inbreeding, the recessive deleterious alleles that cause the inbreeding depression are expected to be removed from the population one after another; especially those that have large negative effects on fertility and viability. As a result, a severe inbreeding event is expected to be more serious for species with large out-crossing populations, compared with species that have a long term history of inbreeding, as for example a self-fertilising plant which previously has undergone a"purging-process" as described above (Lande & Schemske 1985).
Population geneticists have observed that populations may experience reduced fertility and viability if deleterious mutations are accumulated in the gene pool, especially alleles that individually have small deleterious effects. These mutations will never reach high frequencies in large populations with sexual reproduction, where selection will prevent the deleterious alleles from becoming common. However, in small isolated populations where genetic drift may be a stronger force than natural selection, these deleterious alleles could increase in frequency and might even be fixed by chance. The fixation of deleterious alleles in small populations may ultimately affect the survival of these populations, in spite of the fact that each allele individually only has a small effect on population growth. Every fixation of a deleterious mutation will decrease the population size, which in turn enables the fixation of other deleterious mutations – the population has been trapped in a downward spi-ral. In theory, the onset of such a "mutational meltdown" potentially can occur in relatively large populations (Ne > 1000), in which case the extinction will take place after many thousands of generations (Lande 1995).
The risk that different populations will be fixed for exactly the same dele-terious mutations is minute. As a consequence, migrants from different popu-lations will often pass on alleles that lead to higher fitness compared with the deleterious mutations that happen to be fixed in the local gene pool. These “foreign” alleles will be favoured by natural selection, and will increase in frequency in the local gene pool. As a result, this will restore the viability and fertility of the receiving population, a phenomenon termed the "genetic rescue effect" (Ingvarsson 2001; Keller & Waller 2002).
3.2.1 The effects on fertility and viability in natural populations
There are numerous studies investigating if populations with low levels of genetic variation suffer from the loss of genetic variation, inbreeding or