En kunskapssammanställning
ANDERS JARNEMO, WIEBKE NEUMANN, GÖRAN ERICSSON, PETTER KJELLANDER OCH HENRIK ANDRÉN
NAT URV ÅR DSV ERK ET En kunskapssammanställning
Författare: Anders Jarnemo, Wiebke Neumann, Göran Ericsson, Petter Kjellander och Henrik Andrén
Beställningar
Ordertel: 08-505 933 40 E-post: natur@cm.se
Postadress: Arkitektkopia AB, Box 110 93, 161 11 Bromma Internet: www.naturvardsverket.se/publikationer
Naturvårdsverket
Tel: 010-698 10 00 Fax: 010-698 16 00 E-post: registrator@naturvardsverket.se Postadress: Naturvårdsverket, 106 48 Stockholm
Internet: www.naturvardsverket.se ISBN 978-91-620-6819-6
ISSN 0282-7298 © Naturvårdsverket 2018
Förord
Med anledning av förekomsten av avmagringssjuka (”Chronic wasting disease”, CWD) hos vildren, älg och kronvilt i Norge (Tranulis m.fl. 2016; Hansen m.fl. 2017) syftar föreliggande rapport till att sammanställa kunskap om
hjortdjurspopulationer i Sverige, som en del i ett underlag för en handlingsplan under utarbetande av berörda myndigheter (Jordbruksverket, Naturvårdsverket, Statens veterinärmedicinska anstalt (SVA)). På uppdrag av Naturvårdsverket (NV-07973-17) har kunskap om förekomst och fördelning, populationstätheter, biologi, hemområdesstorlekar och rörelsemönster, barriäreffekter samt
mellanartsinteraktioner sammanställts för älg, rådjur, kronvilt och dovvilt.
Rapporten är författad av Anders Jarnemo vid Högskolan i Halmstad och Sveriges lantbruksuniversitet (SLU) samt Wiebke Neumann (SLU), Göran Ericsson (SLU), Petter Kjellander (SLU) och Henrik Andrén (SLU)
Författarna svarar själva för rapportens innehåll. Stockholm i april 2018
Innehåll
FÖRORD 3 SAMMANFATTNING 5 SUMMARY 7
FÖREKOMST AV ÄLG, RÅDJUR, KRONVILT OCH DOVVILT I SVERIGE 9
ÄLG 11
RÅDJUR 17 KRONVILT 22 DOVVILT 28 MELLANARTSINTERAKTIONER ÄLG, RÅDJUR, KRONVILT OCH
DOVVILT 31 KÄLLFÖRTECKNING 33
Sammanfattning
Denna rapport är en kunskapssammanställning över det svenska hjortviltets utbredning, biologi, hemområdesstorlekar, rörelsemönster och
mellanartsinteraktioner. Kunskapssammanställningen är del i ett underlag för en handlingsplan under utarbetande av berörda myndigheter (Jordbruksverket, Naturvårdsverket, Statens veterinärmedicinska anstalt (SVA)) med anledning av risken för spridning av avmagringssjuka (Chronic wasting disease, CWD). Rapporten sammanställdes på uppdrag av Naturvårdsverket (NV-07973-17). Förekomst av älg, rådjur, kronvilt och dovvilt i Sverige har kartlagts med hjälp av statistik över avskjutning och viltolyckor. Älg och rådjur är fördelat över hela landet, där rådjuren förekommer i högre utsträckning söder om den biologiska norrlandsgränsen (Limes norrlandicus, genom södra Värmland i väster till Gästrikland i öster) jämfört med norr om denna. Kronvilt förekommer i samtliga län förutom Norrbottens och Gotlands län, men har en fläckvis utbredning med huvudsaklig utbredning i Götaland och södra Svealand. Starka populationer finns i Skåne, Kalmar län och Sörmland. Dovviltet förekommer främst i södra Sverige med tyngdpunkt i Skåne, Östergötland och Sörmland.
Älgen är en solitärlevande art med undantag av kor med kalvar. Gruppbildning kan dock uppstå på platser med attraktiv föda, speciellt under vintern. Även om
vandringar kan styras av viltstängsel, infrastruktur, vattendrag och topografi, finns inga absoluta barriärer i landskapet för älg. Medan älgar i södra Sverige är mer stationära, är det i Norrland vanligt med säsongsvandringar på upp till 200 km. Hemområdesstorlek kan variera kraftigt mellan olika populationer, men generellt har norrländska älgar större hemområden än de sydsvenska, med maximalvärden 29 000 ha respektive 6 000 ha. Likaså har inlandsälgar större hemområden än kustälgar i Norrland.
Rådjur lever ensamma eller i små familjegrupper. Utvandringsfrekvensen är hög för båda könen. Arten verkar tämligen okänslig för olika typer av barriärer i landskapet och har en snabb spridningskapacitet. I södra Sverige rör det sig vanligen om en genomsnittlig utvandring på 1-3 km, men sträckor på upp till 100 km har observerats på Grimsö. I Norrland är utvandringsfrekvensen mycket hög och här utvandrar rådjuren längre sträckor med i genomsnitt 115-125 km med en längsta observerad utvandring på 470 km. I Syd- och Mellansverige verkar rådjurs hemområden sällan överstiga 100 ha, men man kan förvänta sig större
hemområden i jordbruksdominerade landskap. I Västerbotten har hemområden på upp till 2 000 ha uppmätts.
Kronvilt är grupplevande. Hindarna är trogna sitt uppväxtområde och spridningen går långsamt. Hjortarna är mer benägna att utvandra. Kronvilt är känsliga för barriärer, vilket ytterligare fördröjer spridning. Hindar i Skåne har en genomsnittlig
hemområdesstorlek på 2 600 ha (max. 5 900 ha), medan hindar i Kolmården har 1 200 ha (max 1 800 ha) och hindar på Hunneberg 900 ha (max. 1 500 ha). Medan de mellansvenska hindarna verkar stationära, kan skånska hindar säsongsvandra eller pendla på avstånd upp till 25 km. Hjortarna i Kolmården och i Skåne hade vinter-sommarområden på i genomsnitt 3 400 ha (max. 6 600 ha) respektive 4 300 ha (max. 8 700 ha). Det är vanligt både i Kolmården och i Skåne att hjortarna säsongsvandrar mellan vinter-sommarområde och brunstområde upp till 23 respektive 47 km. Under brunsten kan hjortar också besöka två eller flera brunstlokaler belägna på avstånd upp till 18 km ifrån varandra.
Dovvilt är utpräglat grupplevande och kan bilda mycket täta populationer. Spridningen till nya områden går mycket långsamt. Det verkar vanligt att unga hjortar utvandrar, medan utvandringen av hindar är låg. På Koberg har
hemområden för hindar mätts till i genomsnitt 270 ha (max. 647 ha). För hjortar har hemområden på upp till 1 760 ha uppmätts. Av 18 hjortar gjorde fyra
säsongsvandringar på mer än 5 km mellan brunstplats och vinter-sommarområde. Såväl habitat- som födoöverlapp kan vara stort mellan älg, rådjur, kronvilt och dovvilt. Arterna kan också ses beta i varandras närhet på fält med attraktiva grödor, liksom att de kan utnyttja samma foderplatser och detta ibland samtidigt.
Aggressivt beteende från dovvilt kan dock förmå rådjur och kronvilt att dra sig undan. En negativ effekt av dovvilt på rådjur härrör troligen både från
Summary
This is a review of the biology, distribution, home range sizes, movement patterns, and interspecific interactions of the four wild living cervid species in Sweden. The review is part of background information for an action plan under preparation by concerned authorities (The Swedish Board of Agriculture, The Swedish
Environmental Protection Agency, and the National Veterinary Institute) due to the risk of spreading of Chronic Wasting Disease, CWD. The report was compiled on behalf of The Swedish Environmental Protection Agency (NV-07973-17.
Distributions of moose (Alces alces), roe deer (Capreolus capreolus), red deer (Cervus elaphus), and fallow deer (Dama dama) in Sweden were mapped using statistics on harvest and traffic collisions. Moose and roe deer are spread throughout Sweden, but with a stronger presence of roe deer south of Limes
norrlandicus (approximately south of latitude 60°N). Red deer are established in
all counties except for Norrbotten and Gotland counties, but with large variations in abundance, and a main distribution in Götaland and southern Svealand. The largest populations are found in Skåne, Kalmar and Södermanland counties. Fallow deer mainly occur in southern Sweden with especially high population densities s in the counties of Skåne, Östergötland, and Södermanland.
Moose are generally solitary, but groups can form at places with attractive forage, especially in winter. Although movements are affected by road fences,
infrastructure, topography, lakes, and rivers, there are no absolute barriers in the landscape for moose. Whereas moose in southern Sweden are general stationary, migrations up to 100 km are common in northern Sweden, but migration distances can be up to 200 km. Home range size varies regionally with generally larger home ranges in northern compared to southern Sweden, with maximal values up to 29 000 ha and 6 000 ha respectively. In addition, inland moose generally have larger home ranges than coastal moose in northern parts of the country.
Roe deer are solitary or live in small family groups. Dispersal frequency is high in both sexes. The species appears fairly insensitive to landscape barriers and
population dispersion is rapid. In southern Sweden dispersal distance is on average 1-3 km, but distances up to 100 km have been observed at Grimsö. In northern Sweden, dispersal frequency is very high and dispersal distances are longer with an average of 115-125 km and the furthest observed dispersal distance is 470 km. Home range sizes up to 2 000 ha have been observed in Västerbotten. In Götaland and Svealand, home ranges are in general below 100 ha, although larger sizes may be expected in landscapes dominated by agriculture.
Red deer are group-living. Females are philopatric to their natal area and dispersal is slow. Males are more prone to disperse. Red deer seem sensitive to barriers, which further slows down dispersal. Females in Skåne have an average home range
size of 2 600 ha (max. 5 900 ha), females in Kolmården 1 200 ha (max. 1 800 ha), and females at Hunneberg in Västergötland 900 ha (max. 1 500 ha). Whereas females in Kolmården and Västergötland seem stationary, females in Skåne can migrate or commute up to at least 25 km. Males in Kolmården and Skåne had winter-summer areas of on average 3 400 ha (max. 6 600 ha) and 4 300 ha (max. 8 700 ha) respectively. It is common for males both in Kolmården and Skåne to migrate between their winter-summer area and their rutting area, at distances up to 23 and 47 km respectively. Within the rutting season, males can move up to at least 18 km between different rutting grounds.
Fallow deer are living in mainly gender-segregated groups and can occur in very high densities. Dispersal rate is very slow. It seems common that young males disperse, whereas female dispersal is rare. In Koberg in Västra Götaland county female annual home range sizes were estimated to on average 270 ha (max. 647 ha), whereas male home range sizes can be up to at least 1 760 ha. Out of 18 males, four conducted seasonal migrations between their winter-summer area and their rutting area stretching further than five km.
Habitat and diet overlap between moose, roe deer, red deer, and fallow deer can be high. They can be seen foraging in the vicinity of each other in fields with
attractive crops, and can also use the same artificial feeding sites, sometimes simultaneously. Aggressive behavior from fallow deer can cause roe and red deer to leave. A negative effect of fallow deer on roe deer likely depends both on forage and interference competition.
Förekomst av älg, rådjur, kronvilt
och dovvilt i Sverige
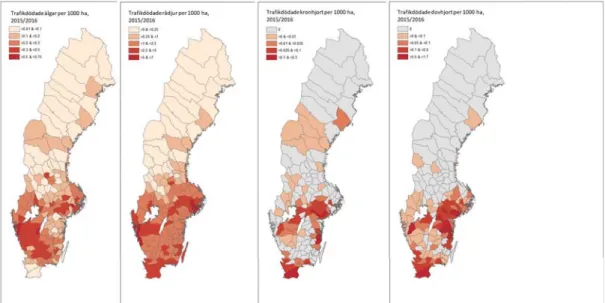
Statistik om avskjutningar (www.viltdata.se) och viltolyckor (www.viltolycka.se) kan användas som ett index för hjortpopulationernas fördelning och förekomst. Dessa data bör dock inte enskilt ses som en skattning av populationstäthet eftersom avskjutning såväl som antal viltolyckor påverkas av fler faktorer än enbart arternas täthet. Avskjutningen speglar förvaltningens målsättning inom respektive område som i sin tur grundas på olika faktorer som lokala intressen och konflikter (t ex. betesskador, viltolyckor, jaktintresse). Antalet viltolyckor påverkas av trafikvolym, infrastrukturens täthet och viltstängsel. Trots dessa begränsningar kan
avskjutnings- och viltolycksstatisk bidra med information om (a) var i landet respektive art förekommer och (b) var arten förekommer i högre eller lägre utsträckning. Viltolycksdata ger dessutom möjlighet att upptäcka arter i nya områden och som under en expansionsfas inte jagas än, men där trots allt några enstaka olyckor med arten förekommer (exempelvis dovvilt i Nordmalings, Edas, Älvdalens och Orsas kommuner, Fig. 2).
Antalet skjutna djur (klövvilt) eller viltolyckor är sammanställt per 1000 ha på älgförvaltningsområden (ÄFO) och resultaten är sammanställda inom
forskningsprogram BEYOND MOOSE (http://www.viltforskning.se/beyond-moose.html), som är finansierat av Naturvårdsverket via medel ur viltvårdsfonden. Älg (Alces alces) och rådjur (Capreolus capreolus) är fördelat över hela landet där rådjuren förekommer i större utsträckning söder om Limes norrlandicus (ungefär söder om breddgraden 60°N) jämfört med sin förekomst norrut (Fig. 1 & 2). Dovvilt (Dama dama) förekommer framför allt i södra delen av landet med starkare förekomst i Sörmland, Östergötland och Skåne (Fig. 1). Kronviltets (Cervus elaphus) fördelning sträcker sig längre norrut och artens förekomst är fläckvis starkare (t.ex. Åre och Nordmaling kommun, Fig. 1). Kronviltets huvudsakliga förekomst är dock söder om Limes norrlandicus med tyngdpunkt i Sörmland, Kalmar och Skåne (Fig. 1). Älg finns i varierande tätheter i hela Sverige, förutom på Gotland.
Figur 1. Avskjutningsdata från jaktsäsongen 2015/2016 för älg, rådjur, kron- och dovvilt på älgförvaltningsnivå (ÄFO). Avskjutningsdata från www.viltdata.se, Jägareförbundet. Resultaten är sammanställda inom forskningsprogram BEYOND MOOSE (http://www.viltforskning.se/beyond-moose.html), som är finansierat av Naturvårdsverket via medel ur viltvårdsfonden.
Figur 2. Fördelning av viltolyckor med älg, rådjur, kron- och dovvilt på älgförvaltningsnivå (ÄFO-nivå) under jaktsäsongen 2015/2016. Baserad på
viltolycksdata från nationella viltolycksrådet (www.viltolycka.se, Jägareförbundet). Resultaten är sammanställda inom forskningsprogram BEYOND MOOSE
(http://www.viltforskning.se/beyond-moose.html), som är finansierat av Naturvårdsverket via medel ur viltvårdsfonden.
Älg
Älgens biologi
Älgen finns i hela barrskogsregionen, i fjällregionen, i skogs- och
jordbrukslandskapet, samt i södra lövskogsregionen. Älgen äter olika växter under olika årstider; blad från lövträd och örter dominerar under sommar, bärris och ljung under vår och höst, och kvistbete vintertid. Beroende på årstid varierar därmed valet av livsmiljö där löv-, bland- och barrskog är älgens dominerande livsmiljöer. Älgen är en solitärt levande art där individer lever var för sig, förutom älgkor med årskalv som rör sig tillsammans. Under vissa delar av året kan älgarna dock samlas i mindre grupper, särskilt under vintertid. Det styrs bland annat av födans
tillgänglighet i landskapet. Nationellt uppskattas att det finns drygt 300 000 älgar i vinterpopulationen (www.jagareforbundet.se). Skandinaviska älgstammar har en mycket låg medelålder och en hög andel älgkor eftersom älgtjurar löper en större risk att dö under jakten (Solberg m.fl. 2000; Ericsson & Wallin 2001; Ericsson m.fl. 2001). Jakt är den huvudsakliga dödsorsaken för en vuxen älg, även i områden där det finns varg eller björn (Sand m.fl. 2006; Swenson m.fl. 2007). Älgen är en långlivad art där kornas livslängd kan uppgå till över 20 år (Ericsson & Wallin 2001). Älgtjurarnas livslängd är betydlig kortare på grund av ett högre jakttryck och ett snabbare åldrande på grund av högre tandslitage (Ericsson m.fl. 2000). Älgen hävdar inget revir och olika älgar kan ha överlappade hemområden. Ett hemområde är ett område där djuren, vistas men vissa delar som utnyttjas mer och andra mindre beroende på fördelning av födoresurser och landskapets
sammansättning. Andelen älgar som tydligt vandrar mellan vinter- och
sommarområden är större i norra delen av landet. Älgtjurar kan förflyttas sig över stora områden under älgkornas brunst. Efter åtta månader dräktighet föder en älgko vanligtvis 1 - 2 kalvar varje år, beroende på hennes kroppskondition, årliga
foderförhållanden och tidigare reproduktion. Under mycket gynnsamma
förehållande kan älgkon föda tre kalvar, men det är ytterst sällsynt. De flesta älgkor blir könsmogna när de är 2 - 4 år gamla. Medelkalvningsdagen ligger i mitten av maj i södra Sverige och i början av juni i norra Sverige som en följd av att älgarnas parningstid (’brunst’) äger rum tidigare söderut.
Barriäreffekter älg
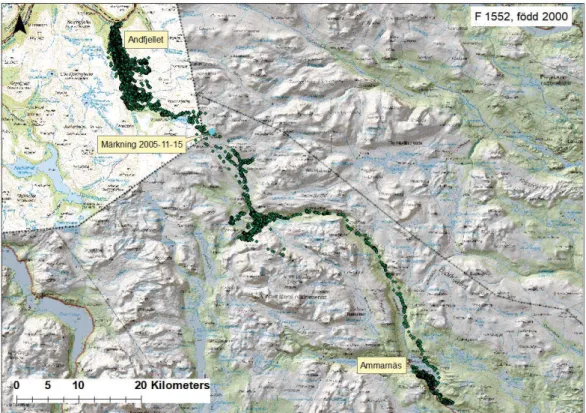
Våra analyser av rörelsedata visar att terrängen påverkar hur vandringsälgar rör sig i landskapet – älgar går framförallt i dalgångar (se exempel av en GPS-märkt älgko som rör sig mellan Sverige och Norge, Fig. 4). Linjära objekt som stora vattendrag leder ofta djurens rörelse (Bartzke m.fl. 2015; Ericsson m.fl. 2017a,b). I områden där tydliga styrande landskapsformationer saknas är vandringen mindre riktad utan älgarna rör sig i alla riktningar (Ericsson m.fl. 2016a).
Figur 3. Rörelse av en älgko märkt inom EU/Interregional IIIA projektet: Älg i MittSkandia. Projektet startade 2004 och är ett samarbete med Norge
(https://www.nina.no/english). Projektet avslutades 2007. Totalt märktes över 100 älgar i tre olika områden för att kartlägga andelen älgar som vandrar från fjällen ner till sina vinterområden i Norge och till det svenska skogslandet.
Generellt kan man säga att födans fördelning i landskapet, klimat, vattendrag, viltstängsel, infrastruktur som vägar med hög trafikvolym (Seiler m.fl. 2005; Neumann m.fl. 2013), samt branter och hög terräng påverkar älgars rörelse, men för en vuxen älg finns inga absoluta barriärer i landskapet. En vuxen älg simmar bra flera kilometer (se exempel förflyttningar mellan öarna i Kalix skärgård, Ericsson m.fl. 2017b), kan passera viltstängsel (Sjöberg 2013), och kan ta sig över relativ hög terräng i fjällen (se exempel Fig. 3). Även om alla dessa faktorer utgör en viss barriär som fördröjer eller påverkar hur snabbt en älg rör sig, kan den ta sig över eller förbi om den vill. Djur följer ofta den energimässigt lättaste vägen genom landskapet och älgen är inget undantag. Barriäreffekterna kan dock vara starkare för kor under den tid korna går med sina kalvar eftersom kalvens mindre kroppsstorlek påverkar förmågan att ta sig över eller förbi ett hinder.
Studieområden älg Sverige
Älgarnas rörelse har följts i 20 olika projekt varav fyra i södra (start 2009) och resten i norra Sverige (start 2003) där älgarna märktes med GPS-halsband
(www.slu.se/alg-forskning). (Ericsson m.fl. 2016a,b; Ericsson m.fl. 2017a,b; Allen m.fl. 2016; Singh m.fl. 2012).
Studieområden i norra Sverige (Västerbotten, Norrbotten) finns i fjällregionen, i fjällbjörkskogsregion och i norra barrskogsregionen (Tabell 1). I områden där älgar visar ett utpräglat vandringsbeteende kan de röra sig över tre vegetationszoner (t ex. områden Arjeplog och Hemavan, Fig. 4A & 4B). Skog dominerar framför allt i områden som ligger i det norra inlandet (>60%). Inslaget av myrar kan vara högt (>25%) i på vissa håll. Dominerande trädslag är tall (Pinus sylvestris) med inslag av contortatall (Pinus contorta) och gran (Pieca abies). Lövträd som förekommer är björk (Betula sp.), asp (Populus tremula) och sälg (Salix sp).
Studieområden i södra Sverige (söder om breddgraden 60°N) ligger i den södra barrskogsregionen, med undantag för Grimsö som tillhör den sydliga zonen av den norra barrskogsregionen (Tabell 1). Studieområdena är betydligt mindre jämfört med områdena i norra delen av landet på grund av att älgarna rör sig över en betydlig mindre yta. Studieområdena i södra Sverige tillhör ett skogs- och jordbrukslandskap där skogsområdena är mindre och mindre sammanhängande jämfört med norra Sverige. Tall och gran är dominerande träslag, men bestånd av ädellövträd förekommer. Studieområdet på Öland intar en särskild roll med många ovanliga livsmiljöer för älg; jordbruksmarker dominerar landskapet och skog förekommer bara i begränsade bestånd medan buskar och sparsam vegetation framför allt på Stora Alvaret, utgör stora delar av landskapet (Tabell 1).
Rörelsemönster och hemområdesstorlekar hos älg
Sammanfattningsvis är älgarna i södra Sverige (söder om breddgrad 60°N) i stor utsträckning stationära (Fig. 4 C), medan i norra Sverige är andelen av älgar som vandrar säsongsmässigt större. I norra Sverige ökar vandringsavståndet från kust till inland (Fig. 4A & 4B). Undantag är studiepopulationen Nikkaluokta (breddgrad 67°N), där andelen av stationära älgar är hög jämfört med andra områden. Älgarna vid Nikkaluokta utmärker sig också genom att de förflyttar sig över relativ liten yta jämfört med älgar i andra områden på samma breddgrad. Istället förflyttar sig älgarna i höjd mellan sommar och vinter (s.k. ”altitudinal migration”). I varje population (söder och norr) ser vi en stor variation i vilken yta en enskild älg rör sig över; vissa älgar har ganska begränsande hemområden medan andra rör sig över större ytor.
Här redovisar vi storlek av områden älgar rör sig över under ett år, här efter kallat hemområden. Hemområden för älgkor i norra Norrland varierar mycket, beroende
på om älgkon tillhör en population vid kusten eller i inlandet (Fig. 4A).
Genomsnittliga hemområden kan vara så små som 1 880 ha (kust) eller så stora som 9 580 ha (Arvidsjaur), och maximala hemområdesstorlek varierar motsvarande mellan populationer (4 960 ha – 23 340 ha; Fig. 4A). Jämfört med älgkorna, uppvisar älgtjurarna ofta andra rörelsemönster där de förflyttar sig över en större yta än korna. Liksom för älgkorna, varierar den genomsnittliga storleken av hemområden för älgtjurar beroende var i Norrbotten älgen är; 2 150 ha (kust) – 14 450 ha (Fig. 4A) och har maximalt uppmätts till 29 120 ha (Arjeplog; Fig. 4A). Samma mönster finns i Västerbotten – ju närmare fjällen, desto större andel vandringsälgar och längre vandringsavstånd (Fig. 4B). Älgar i den svenska fjällkedjan omfattar både ”norska” och ”svenska” vandringsälgar (projektet ”Älg i MittSkandia”, Fig. 4B).
I södra Sverige omfattar ett hemområde av en älgtjur i genomsnitt 2 670 ha eller mindre och kan maximalt vara så stor som 6 100 ha. Älgkornas hemområden är i genomsnitt 1 600 ha och maximal upp mot 4 390 ha (Fig. 4C).
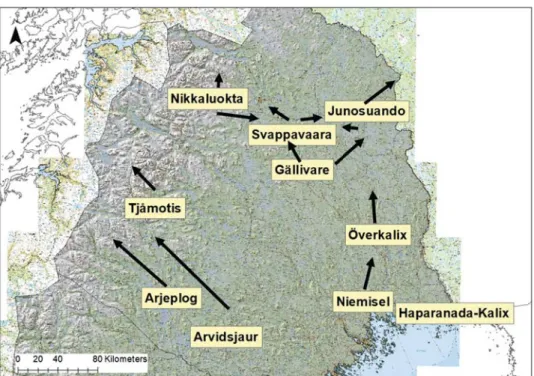
Figur 4. Fördelning av studieområden med GPS-märkta klövviltsarter i tre olika delar av Sverige (A: Norrbotten, B: Västerbotten, C: Södra Sverige).
Figur 4A. Norrbotten
Figur 4C. Fördelning av studieområden med GPS-märkta klövviltsarter i tre olika delar av Sverige (A: Norrbotten, B: Västerbotten, C: Södra Sverige, söder om breddgrad 60°N) för älg (gul, n = 20), rådjur (grön, n = 7), kronvilt (röd, n = 3) och dovvilt (blå, n = 1). Pilar i figur A och B indikerar älgarnas huvudsakliga
Rådjur
Rådjurets biologi
Rådjur har en vid utbredning i Europa. Arten är flexibel vad gäller habitatkrav och hittas i en mängd olika habitattyper (Linnell m.fl. 1998a), men tros ha utvecklats i och anpassats till skogsmiljöer i tidiga successionsstadier (Liberg & Wahlström 1995). Rådjuret är en selektiv kvalitetsbetare som söker de mest näringsrika växtdelarna. Födovalet utgörs av ett brett spektrum växtarter, men rådjuret föredrar örter, blad och knoppar (Duncan m.fl. 1998).
Parningen sker i juli-augusti, men implantationen är fördröjd med fem månader (Sempéré m.fl. 1998). I maj-juni föds 1-3 kid. Rådjur är en typisk ”gömmare” (Espmark 1969; Lent 1974) och kiden ligger dolda och åtskilda från modern det mesta av tiden upp till cirka åtta veckors ålder (Linnell 1994). Getterna föder i regel sina första kid vid två års ålder. Livslängden för rådjur i Skandinavien är upp till 10-12 år, där geten i allmänhet blir något år äldre än bocken.
Rådjur har en hög utvandringsfrekvens för båda könen (Wahlström & Liberg 1995a, Wahlström & Liberg 1995b; Linnell m.fl. 1998b). I april klipper de snart ettåriga fjolårskiden bandet till modern och blir betydligt mer rörliga och företar under vår och sommar exkursioner utanför moderns hemområde. Både getter och bockar, men i högre utsträckning getter, kan återförenas med sin mor under höst-vinter när större grupper bildas (Bideau m.fl. 1987; Wahlström & Liberg 1995a; Linnell m.fl. 1998). De djur som utvandrar gör det i regel under vår-sommar vid ett års ålder. Såväl bockar som getter kan dock utvandra vid två-tre års ålder (Linnell m.fl. 1998). Wahlström & Liberg (1995a) fann att 40 % av de getter som utvandrat (d.v.s. lämnat moderns hemområde) på sommaren återvände till sitt uppväxtområde inför vintern. Denna säsongsmigration kan pågå en eller två vintrar varefter de stannade permanent i området de utvandrat till som ettåringar (Wahlström & Liberg 1995a; Linnell m.fl. 1998).
Getternas utvandring verkar vara frivillig och i stor utsträckning styras av tillgång till resurser. De verkar känsliga för skillnader i kvalitet mellan områden och fördelar sig till synes optimalt i förhållande till resurstillgång enligt ett idealt fritt fördelningsmönster (Wahlström & Kjellander 1995; Linnell m.fl.1998). För unga bockar verkar aggression från revirhävdande bockar vara en viktig faktor som driver på utvandring. Välväxta och välutvecklade ungbockar utvandrar i högre grad och dessa har också visat sig vara mer utsatta för aggressioner från dominanta bockar (Strandgaard 1972; Wahlström 1994; Linnell m.fl. 1998).
Wahlström & Liberg (1995a) fann den högsta utvandringsfrekvensen vid
intermediära tätheter, s.k. ”presaturation dispersers”. De utvandrande djuren var i genomsnitt tyngre än stannande djur och det är möjligt att rådjuren måste uppnå en
viss kondition för att kunna utvandra. Det kan vara så att vid en låg täthet och låg resurskonkurrens förblir fler djur i uppväxtområdet. Med en ökande täthet och konkurrens ökar utvandringsfrekvensen, men vid en hög täthet och hög konkurrens om resurser kan det vara så att många djur är i för dålig kondition för att klara av att utvandra (Wahlström & Liberg 1995a).
Rådjur lever ensamma eller i små familjegrupper på 2 - 3 djur som ofta består av en get och hennes kid samt ibland även en vuxen bock (Hewison m.fl. 1998).
Ettåringar kan ses i sällskap med en vuxen bock eller en familjegrupp strax innan utvandring. Bockar lever mer ensamma än getter under sommar och revirhävdande. Getter som strax ska föda söker avskildhet.
Bockar inrättar på våren (mars) ett revir som upprätthålls över brunsten i juli och augusti (Johansson 1996; Liberg m.fl. 1998). Reviret försvaras genom
revirmarkeringar och genom att aggressivt jaga bort andra bockar (Bramely 1970; Johansson 1996). Det första reviret etableras vid 2 - 4 års ålder (Bramely 1970; Strandgaard 1972; Liberg m.fl. 1998) och bockarna håller sina revir, relativt ortstroget i genomsnitt upp till cirka 5 - 7 års ålder (Johansson 1996; Hewison m.fl. 1998; Liberg m.fl. 1998). Ej revirhävdande bockar kan ha större hemområden och röra sig över flera revirbockars revir. En annan strategi är att hålla sig perifert i små områden om ett par eller några hektar i gränsområdet mellan dominanta bockars revir. Ytterligare en strategi är att inta en underordnad ställning och då tolereras i en dominant bocks revir (Hewison m.fl. 1998).
Getter hävdar inte revir, även om aggressioner mellan getter kan ses. Normalt ses stora överlapp mellan getters hemområden, även om det ofta är fråga om
närbesläktade getter (Liberg m.fl. 1998). Getter har också ett kärnområde i hemområdet som ofta är större än motsvarande hos bockarna och getter kan röra sig över flera bockars revir. Det är vanligt att getter gör ”brunstexkursioner” utanför sitt normala hemområde (Debeffe m.fl. 2014). Bockars och getters
rumsliga utnyttjande är i regel stabilt, varför samma individer av bockar och getter troligen har en nära rumslig relation under en stor del av livet (Hewison m.fl. 1998).
När revirhävdandet avtar under hösten börjar det bli vanligare med grupper på fyra eller fler djur (Stüwe & Hendrichs 1984; Maublanc m.fl. 1987; Danilkin &
Hewison 1996), och det är också vanligare att djur håller sig tillsammans med ett eller flera djur än att de går ensamma (Bramely 1970; Hewison m.fl. 1998). Genomsnittlig gruppstorlek ökar med graden av öppenhet i landskapet (Gerard m.fl. 1995) och i jordbruksdominerade landskap ökar gruppstorlek med storlek på fält och med ökad homogenitet i landskapet (Bresinski 1982). I Skåne är det inte
bryts de större grupperna upp då bockarna också inrättar revir och ungdjur börjar vandra ut.
Barriäreffekter rådjur
Efter att bara ha funnits i Skåne på 1860-talet, lyckades rådjuret under de närmsta 60 åren sprida sig upp till Norrlands kustland, Dalarna, Värmland och in i Norge (Cederlund & Liberg 1995). Med en hög grad av utvandring för båda könen (Wahlström & Liberg 1995a, Wahlström & Liberg 1995b; Linnell m.fl. 1998b; Gaillard m.fl. 2008) och en visad snabb förmåga att kolonisera verkar rådjur tämligen okänsliga för olika typer av barriärer, även om en kombination av större vägar och vattendrag har visat sig minska genflöde mellan områden (Coulon m.fl. 2006). Vattendrag utgör dock inte ett absolut hinder, och båda könen kan
regelbundet och oberoende av säsong simma över stora vattendrag (Toth m.fl. 2016). Vad gäller viltpassager vid vägar, nyttjar rådjur såväl över- som
underpassager (Olbrich 1984; Georgii m.fl. 2007). I fragmenterade landskap verkar spridningen vara starkt förknippad med och gynnas av förekomsten av skog
(Coulon m.fl. 2004).
Studieområden rådjur Sverige
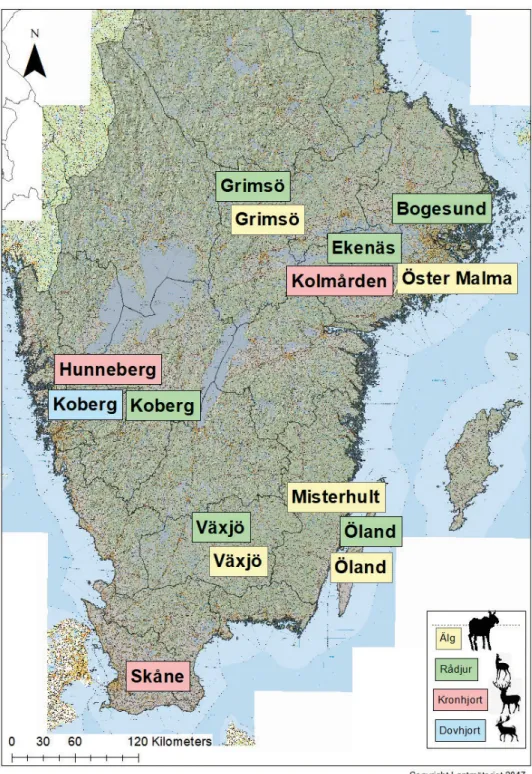
Koberg (58°11´N, 12°41´E, boreonemoral zon) ligger i Västergötland cirka 12 km syd-sydost om Trollhättan. Terrängen är låglänt och med 79 % skog (i huvudsak gran och tall), 16 % jordbruksmark och 2 % myr. Åkermarken används i stor utsträckning för odling av vall för viltbete. På Koberg har dovvilt och rådjur studerats sedan 2006.
Ekenäs (58°58´N, 16°35´E, boreonemoral zon) är beläget i Södermanland, cirka 10 km söder om Flen. Landskapet är ett blandat skogs- och jordbrukslandskap med 65 % skog och 35 % jordbruksmark. Tall och gran dominerar, men björk, klibbal, asp och ek är vanligt förekommande. På Ekenäs studerades rådjur under 1986 - 1999. Bogesund (59°23´N, 18°15´E, boreonemoral zon) ligger i Uppland cirka 10 km norr om Stockholm och strax väster om Vaxholm. Studieområdet består av 65 % skog, 25 % jordbruksmark och 10 % myr, hällmark etc. Tall och gran dominerar. Vanliga lövträd är asp, ask, björk, ek och klibbal. Rådjur har studerats med varierande intensitet på Bogesund sedan 1988.
Grimsö (59°84´N, 15°48´E, södra boreala zonen) ligger i Västmanland mellan Lindesberg och Skinnskatteberg. Området är flackt och domineras av skog (75 %) med inslag av myr (12 %), vattendrag (7 %) och övriga markslag (6 %). Gran och tall är dominerande trädslag med inslag av björk, asp, rönn och sälg.
I Västerbotten (mellanboreal zon) märktes rådjur vid Gunnarn (65°00´N, 17°40E´) och vid Åskilje (64°54´N, 17°52´E) 27 respektive 39 km sydost om Storuman. Landskapet domineras helt av barrskog. Endast i dalgångarna i närheten av bebyggelse finns jordbruksmark (täcker mindre än 1 % av arealen).
Rörelsemönster och hemområdesstorlek hos rådjur
På Ekenäs utvandrade 64 % av bockarna och 56 % av getterna från
uppväxtområdet. Genomsnittlig sträcka var 7,6 km med en längsta sträcka på 40 och 58 km för bock respektive get (Wahlström & Liberg 1995a, 1995b).
Wahlström & Liberg (1995a, 1995b) fann att på Bogesund utvandrade 32 % av bockarna och 41 % av getterna. I genomsnitt gick de iväg 2,7 km. Längsta vandring noterades för en bock som vandrat iväg 10 km när kontakten med
sändaren upphörde. En senare studie fann en genomsnittlig utvandring på 1 km och en längsta på 8 km (Gaillard m.fl. 2008). Av 9 GPS-märkta kid på Koberg
noterades endast ett par längre utvandringar om 12 - 15km, övriga yngre djurs rörelser mättes till enstaka kilometrar (Kjellander opubl.). I Västerbotten
utvandrade 19 av 20 djur, d.v.s. 95 %. Här vandrade dessutom rådjuren betydligt längre sträckor med i genomsnitt 125 km för Gunnarn och 115 km för Åskilje. En get gjorde en vandring som sträckte sig över hela 470 km (Cederlund & Liberg 1995; Wahlström & Liberg 1995a, 1995b). Även på Grimsö utvandrar merparten av ettåringarna. De längsta utvandringssträckorna är ca 100 km medan det vanligen endast rör sig om 2-3km i genomsnittlig utvandringssträcka (Kjellander m.fl., opubl.)
För de mellansvenska områdena är hemområdesstorlekarna tämligen likvärdiga och verkar sällan överstiga 100 ha. Rådjuren på Ekenäs har minst hemområden med 20 - 40 ha för helår och 15 - 30 ha för enskilda årstider (Cederlund & Liberg 1995). På Bogesund anges varierande hemområdesstorlekar beroende på kön, årstid och populationstäthet med större hemområden vintertid och vid lägre täthet (Kjellander m.fl. 2004). Bockarnas sommarhemområden minskas från 50 ha till 35 ha vid ökande täthet, medan vinterhemområdena går ner från drygt 100 ha till cirka 40 ha. Sandhammar (2005), visade dock att medelstorleken på bockarnas revir på
Bogesund under den mest intensiva brunsten i juli-augusti var 23,5 ha. Getternas sommarhemområden ligger på 25 - 40 ha och vinterhemområdena på 35 - 70 ha. Guillet m.fl. (1996) fann också likvärdiga vinterhemområden för getter på Bogesund och Grimsö med 57 respektive 59 ha, medan Öhman (2017) redovisar sommarhemområden för getter på 136 ha på Grimsö och 65 ha på Bogesund, medan Morellet m.fl. (2013) fann hemområden för getter på 68 ha på Koberg.
600 ha. Säsongsmigrerande rådjur har vinterhemområden på 50 - 100 ha, men har sommarhemområden på upp till 2 000 ha.
Det finns inte svenska data på hemområdesstorlek för rådjur i jordbruksdominerade landskap. Utländska studier tyder dock på att vinterhemområden i öppna landskap kan öka i storlek där föda och skydd är mer utspritt. Vinterhemområden för vuxna rådjur kan då ligga på 400 - 500 ha, och för juvenila cirka 700 ha (Danilkin & Hewison 1996; Hewison m.fl. 1998).
Kronvilt
Kronviltets biologi
Kronvilt är en art som är anpassad till starka konkurrensförhållanden i en stabil och förutsägbar miljö (Liberg & Wahlström 1995). Arten har troligen utvecklats i ett blandlandskap av lövdominerade skogar och öppen terräng. De är relativt
långlivade och hindar kan uppnå en ålder av mer än 20 år och fortfarande ändå vara fertila. De får en kalv och mindre än 1 % som föder två kalvar (Mitchell & Lincoln 1973; Langvatn m.fl. 1996; Jarnemo 2014). Hjortarna har en sen mognad och blir inte kroppsligt fullvuxna förrän vid sex till åtta års ålder och är som starkast avseende dominans under brunsten samt hornstorlek vid 10 till 14 års ålder. Hjortar i det vilda torde sällan bli äldre än cirka 15 år. Kronvilt är en blandbetare där en stor del av födan hämtas i mark- och fältskikt i form av gräs, örter, bärris, ljung och lav, men där även löv, knoppar, bark och frukt kan ingå (Lavsund 1976; Gebert & Verheyden 2001).
Hindar är trogna sitt uppväxtområde (filopatriska) och spridning verkar snarare ske genom att gränsen för hemområdet successivt flyttas, än att hindar utvandrar (Clutton-Brock m.fl. 1982; Catchpole m.fl. 2004). Därmed får man en
jämförelsevis långsam spridning av kronvilt. Utvandring av hjortar är vanligare (Clutton-Brock m.fl. 1982; Catchpole m.fl. 2004) och genetiska studier bekräftar bilden av kronvilt som en art där hjortarna utvandrar och hindarna är filopatriska (Nussey m.fl. 2005; Frantz m.fl. 2008; Haanes m.fl. 2011). Ofta verkar det vara hjortar som först dyker upp och uppehåller sig i nya områden utanför brunsttid. Könssegregation, som kan resultera i att hjortarna säsongsvandrar mellan brunstområden och vinter-sommarområden (Jarnemo 2008) samt förflyttningar mellan olika brunstområden (Jarnemo 2011), är processer som kan tänkas leda till en högre spridning för hjortar än för hindar.
Ettåriga hindar lämnar modern i samband med kalvning, men kontakten återupptas och döttrar etablerar i regel ett hemområde som till stor del överlappar med
moderns, och rör sig också ofta i samma grupperingar som modern. Ettåriga hjortar har ett lösare band till modern och i samband med kalvning minskar kontakten kraftigt och för hjortar äldre än två år är såväl överlapp med moderns kärnområde som kontakt med modern minimal (Clutton-Brock m.fl. 1982).
Kronvilt är en art som uppvisar s.k. könssegregation (Clutton-Brock m.fl. 1982, 1987; Bonenfant m.fl. 2004). Större delen av året lever hindar och hjortar åtskilda i olika grupper och ofta också i olika områden (Jarnemo 2008). Hindarna lever i grupper som består av närbesläktade hindar, deras kalvar och fjolårskalvar. Även
Grupper med hjortar, s.k. herrklubbar eller ungkarlsflockar, är än mer löst sammansatta än hindgrupperna. Vilka hjortar som ingår i ungkarlsflockarna kan fluktuera kontinuerligt och ändras nästan timme för timme då hjortar lämnar eller ansluter. Släktskapet mellan ingående hjortar är också mindre jämfört med i hindgrupper. Ofta kan en åldersuppdelning observeras både inom och mellan ungkarlsflockar med yngre hjortar som håller samman och äldre hjortar som tyr sig till varandra. Riktigt gamla hjortar kan dock ofta ses ensamma (Clutton-Brock m.fl. 1982).
Kronvilt är polygyna (d.v.s. en hjort betäcker flera hindar) och har vanligen ett brunstsystem med haremsbildning. De starkaste hjortarna samlar och försöker hålla ihop ett harem av hindar som försvaras aggressivt mot andra hjortar. I en naturlig stam med någorlunda jämn könsfördelning och med en naturlig tillgång på fullvuxna hjortar (d.v.s. hjortar som är från 6-8 år till 12-14 år gamla), är det först när hjortarna blir fullvuxna i kroppen, d.v.s. i åldern sex till åtta år, som de når framgång under brunsten och lyckas bli haremshållare s.k. platshjortar. Hjortar yngre än sex år är i princip att betrakta som tonåringar och dessa har i regel små chanser att hävda sig under brunsten, förutsatt att tillgången på vuxna hjortar är tillfredsställande (Clutton-Brock m.fl. 1982). Yngre hjortar eller vuxna hjortar som inte vågar utmana platshjorten håller sig i utkanterna av platshjortens harem och kan också besöka flera brunstlokaler i jakt på hindar (Jarnemo 2011). En alternativ strategi för vuxna hjortar som inte är platshjortar, kan också vara att ställa sig på en plats och genom bröl försöka locka till sig hindar.
Det intensiva brölet är dels en styrkedemonstration gentemot andra hjortar, men också ett sätt att attrahera hindar. Genom att lyssna på brölet kan hindarna bedöma hjortens styrka och kondition (McComb 1991, Reby m.fl. 2001, Reby & McComb 2003). Man har även kunnat visa att igångsättningen av hindarnas brunst gynnas av att de hör mycket bröl (McComb 1987). En tidigare brunst innebär en tidigare betäckning, vilket i sin tur gör att kalvarna föds tidigare och ges mer tid att förbereda sig inför sin första vinter och därmed troligen får en bättre start i livet. Man har sett att kronhindar söker sig till vuxna hjortars harem och observationer tyder på att de i det längsta försöker undvika att para sig med hjortar som är yngre än fem år (Clutton-Brock m.fl. 1982). Brist på fullvuxna hjortar kan därmed leda till ombrunst och sena betäckningar. Förutom chansen till en starkare avkomma, slipper hindarna i skydd av en stark platshjort stressande uppvaktning från yngre hjortar och kan på så sätt spara energi och söka föda i lugn och ro, vilket därmed ger en bättre förberedelse inför vintern och den kommande dräktigheten. Yngre hjortar brölar mycket litet och i en population med en låg andel hjortar och en låg medelålder för hjortarna, är brölaktiviteten mycket låg, vilket då kan resultera i att brunsten kommer igång senare med senare betäckningar som följd. Sammantaget innebär detta att de vuxna hjortarna har mycket stor betydelse i kronviltets rörelser, sociala organisation och ekologi.
Barriäreffekter kronvilt
Kronvilt har visat sig känslig för olika typer av barriärer och hinder i landskapet i form av bland annat infrastruktur och större vattendrag (Patthey 2003, Pérez-Espona m.fl. 2008; Frantz m.fl. 2012). Det verkar som att kronvilt i sin utbredning kan göra halt under flera år vid större vägar och vattendrag med följden att man kan ha en hög täthet på ena sidan och få eller inga djur på andra sidan. När sedan djur bestämmer sig för att korsa vägen eller vattendraget, kan man uppleva en snabbare spridning på andra sidan (A. Jarnemo pers. obs.). Kronvilt verkar mindre benägna att använda underpassager vid vägar jämfört med andra viltarter (Olbrich 1984; Georgii m.fl. 2007; Frantz m.fl. 2012). Kronviltets känslighet för barriärer har visat sig kunna leda till isolerade populationer, i förlängningen med
inavelseffekter som följd (Kuhn m.fl. 2003; Zachos m.fl. 2007).
Studieområden kronvilt Sverige
Skåne (55°4´N, 13°40´E, nemoral zon). Flackt låglandsområde med inslag av böljande kullar och åsar. Landskapet domineras av jordbruk blandat med skog av gran tall, lärk, bok, björk, ek och ask. Här finns data från 12 hindar och 4 hjortar som märkts i södra-centrala Skåne.
Hunneberg (58°20´N, 12°28´E), boreonemoral zon) är ett platåberg vid Vänerns södra strand, strax öster om Vänersborg och Trollhättan i Västergötland. Berget avgränsas av branta stup gentemot omgivande jordbruksmark. Toppen, som når 90 m över omgivningen, är flack och täcks av barrskog på mager mark med inslag av myrar, ekhagar och sjöar. Här finns data från 4 hindar.
Kolmården (58°48´N, 16°24´E, boreonemoral zon) i Östergötland-Södermanland. Här har kronvilt försetts med GPS-halsband på fyra olika marker: Stavsjö, Virå Bruk, Valinge och Ericsberg. Den västra delen av studieområdet (Stavsjö och Virå) utgörs ett homogent barrskogslandskap med 84 % skog, 8 % myr, 6 % hällmark och 1 % jordbruksmark. Inslaget av jordbruksmark ökar i östra delen (Valinge) där 64 % är tall- och grandominerad skog, 2 % myr, 8 % hällmark och 22 %
jordbruksmark. Från Kolmården finns data från 12 hindar och 10 hjortar.
Rörelsemönster och hemområdesstorlek hos kronvilt
Det finns inte bra svenska data på utvandringsfrekvens eller spridningssträckor hos kronvilt. Av 14 öronmärkta kalvar i forskningsprojektet i Kolmården, finns ett,
ifrån märkplatsen. Det finns dock observationer i Sverige av kronvilt i områden där det inte finns etablerade populationer, vilket kan ge en antydan om
spridningskapacitet. I Lavsunds (1975) redogörelse för utbredning och
observationer av kronvilt i Sverige under 1900 - 1973, finns flera observationer av enstaka individer ända upp i Lappland, långt från hägn och etablerade populationer. Hägn var ovanligt i Norrland under denna period och Lavsund finner det troligt att de flesta av dessa norrländska observationer rör djur som vandrat från Norge. Ett exempel är ett handjur som uppträdde i Vittangi 1955 och som då befann sig cirka 550 km från närmsta norska förekomst. Även om till exempel dagens populationer i Jämtland och Värmland har sin grund i djur med härkomst från hägn (Sennstam 2005, Höglund m.fl. 2013), kan det samtidigt förekomma långvandrare från Norge. Det finns exempelvis tecken på att det kan ha tillkommit norsk inblandning i kronviltpopulationen på Hunneberg (Höglund m.fl. 2013). Det finns även uppgifter om att hindar märkta i Norge vid Askim, ska ha observerats på svenska sidan vid Sunne i Värmland, en sträcka på cirka 110 km. Observationerna tyder på att hindarna gör en säsongsvandring mellan Askim och Sunne.
Även vad gäller utländska studier är tillgången på utvandringsdata bristfällig. Clutton-Brock m.fl. (1982) kunde konstatera att en tredjedel av hjortkalvarna hade lämnat studieområdet senast före sin fjärde vinter. Då studieområdet här ligger på en ö, är det svårt att dra några slutsatser om spridningssträckor. Det finns dock uppgifter från Skottland att hindar märkta som kalvar, i vuxen ålder har påträffats upptill 31 km från födelseplatsen (Daniels och McClean 2002). En belgisk studie (Prévot & Licoppe 2013) rapporterar en spridningssträcka på i genomsnitt 1,9 km (max 6,3 km) för hindar och 4,8 km (max 16,8 km) för hjortar.
De skånska hindarna har en genomsnittlig hemområdesstorlek på 2 600 ha (max 5 910 ha), att jämföra med knappt 1 200 ha (max 1 824 ha) för Kolmårdenhindarna och knappt 900 ha (max 1 545 ha) för Hunnebergshindarna. Hjortarna har större hemområden än hindarna, och med förbehållet att datamaterialet är litet, så har de skånska hjortarna större hemområden än Kolmårdenhjortarna med i genomsnitt 4 300 ha (max 8 700 ha) respektive 3 400 ha (max 6 600 ha).
Resultaten tyder på att i ett mer öppet, fragmenterat och jordbrukspåverkat landskap, som exempelvis Skåne, kan man förvänta sig en större
hemområdesstorlek än i ett homogent skogslandskap, som exempelvis i
Kolmården, något som också påvisats för kronvilt i andra studier (Bevanda m.fl. 2015). Det positiva sambandet mellan andel öppen mark och hemområdesstorlek sågs dels i jämförelsen mellan Skåne och de mellansvenska områdena, dels inom respektive studieområde.
Hos kronvilt lever vanligen hindar och hjortar åtskilda utanför brunstperioden (Clutton-Brock m.fl. 1982, 1987; Bonenfant m.fl. 2004). Hjortarna lämnar ofta brunstplatserna efter brunsten och vandrar till särskilda områden där de tillbringar resten av året (Jarnemo 2008). En sammanställning av kartlagda vandringar av 96
olika hjortar mellan brunstområde och sommar-/vinterområde i Skåne under 1969 - 2007, visade att genomsnittligt avstånd mellan brunstobservationer och sommar-/vinterobservationer var 13,9 km med en längsta observerad vandring på 47 km (Jarnemo 2008; Jarnemo 2014).
Av de 10 Kolmårdenhjortarna med GPS-halsband gjorde sju en vandring mellan brunstområde och sommar-/vinterområde. Tre hjortar hade brunstområden som helt överlappades av sommar-/vinterområdet. För de hjortar som uppvisade en
vandring, var det genomsnittliga avståndet för förflyttningen (centrum till centrum) 10,6 km (min 5,6 km, max 18,5 km) alternativt 15,0 km (min 8,3 km, max 23,5 km) mätt som avståndet mellan de från varandra längst belägna punkterna (Jarnemo 2014).
En skånsk hind gjorde en säsongsvandring på 10 km och två hindar vandrade mellan två respektive tre olika delområden på avstånd upp till 25 km. Motsvarande säsongsvandringar eller vandringar mellan områden observerades inte för hindar i Kolmården eller på Hunneberg där hindarna var stationära i sina
märkningsområden. En av Kolmårdenhindarna verkade dock lämna sitt
huvudområde i ett blandat skogs- och jordbrukslandskap i samband med strängare vinterförhållanden, och då istället söka sig till ett mer homogent barrskogsområde. De svenska studierna tyder således på att hindar i jordbrukslandskap inte bara har större hemområden än hindar i skogsdominerade landskap, de verkar också mer benägna att vandra mellan olika delområden jämfört med hindar i skogslandskap som är mer stationära.
Genom använda individuellt identifierade (med hjälp av hornkaraktärer) hjortar, studerades hjortars förflyttningar mellan olika brunstlokaler under en och samma brunstsäsong (Jarnemo 2011). Under 1998-2009 noterades förflyttningar av hjortar mellan olika brunstlokaler vid 91 tillfällen. Den längsta observerade sträckan som en hjort tillryggalade mellan två olika brunstlokaler var 18,5 km (genomsnitt 8,1 km). Av de 91 vandringarna var 7 gjorda av ettåringar, 46 av unghjortar (bedömd ålder 2 - 5 år) och 38 av vuxna hjortar (bedömda som sex år och äldre). Alla brunstlokaler inom inventeringsområdet var länkade till minst två andra
brunstlokaler genom hjortarnas vandringar. För alla hjortar kombinerat, liksom för ettåringar och unghjortar, var antalet hindar på den andra besökta brunstlokalen högre än på den första lokalen. Likaså var kvoten hindar/vuxna hjortar högre på den andra lokalen än på den första. Slutsatsen blir att de observerade
förflyttningarna är relaterade till intra-sexuell konkurrens och genomförda för att förbättra chanserna till reproduktionsframgång. Resultaten avslöjar ett betydande utbyte av hjortar brunstlokaler emellan under brunsten, vilket tyder på en potential för sekundär spridning hos kronvilt och på en stor kapacitet av genflöde mellan
att kronhindar kunde göra brunstexkursioner på upp till 4 km utanför hemområdet. Även för svenska kronhindar har brunstexkursioner på cirka 4 km observerats (A. Jarnemo, opubl.)
Kronviltet är mycket känsligt för olika typer av störningar, vilket gäller såväl jakt som militära aktiviteter och friluftsliv (Jeppesen 1987; Bateson & Bradshaw 1997; Phillips & Alldredge 2000; Sunde m.fl. 2009; Sibbald m.fl. 2011). Störningen kan få kronvilt att lämna sina hemområden, och en ökad omfattning av störning kan också generera större hemområden (Jeppesen 1987; Sunde m.fl. 2009). I samband med drevjakter på kronvilt i Kolmården uppmätte man flyktsträckor på mellan 0,4 och 15 km (medel 2,5 km; Jarnemo 2014; Jarnemo & Wikenros 2014). I
genomsnitt återkom de till hemområdena efter 23 timmar (2 - 88 timmar). För hjortarna i Kolmården observerades åtta flykter. Genomsnittlig flyktsräcka var 5,1 km (1,2 - 13,3 km). Tre av flykterna genomfördes till fullo under pågående jakt, tre under natten efter jakt och vid två flykter gjordes en första del under jakt och en andra del under natten efter jakt. Vid tre av flykterna (3 olika individer) i början av jaktsäsongen lämnade hjortarna sin brunstplats och gick till sitt
vinter-sommarområde, för att återvända först vid nästa års brunst. Samtliga hjortar som gjorde en brunstvandring hade lämnat brunstområdena senast en vecka efter jaktpremiären och återvänt till sommar-/vinterområdet. Den höga jaktliga
aktiviteten första jaktveckan verkar trigga hjortarna att lämna brunstområdena och återvända till vinter-sommarområdet.
Dovvilt
Dovviltets biologi
Dovvilt hade en naturlig utbredning i kontinentala Europa ända upp till Danmark, fram till för 10 000 – 50 000 år sedan, då den verkar ha dött ut åtminstone i Centraleuropa. Helt säkert är dock att arten överlevde öster om Medelhavet. Dovvilt är lätt att hålla i fångenskap och åtminstone sedan romartiden har djur transporterats över stora delar av Europa (Chapman & Chapman 1975). Till Sverige introducerades dovvilt på 1500-talet och kom att hållas i hägn på kungliga marker och vid adelns slott. Dessa tidiga s.k. djurgårdar och mer sentida hägn har kommit att fungera som spridningscentra för arten i Sverige. Frilevande
populationer började uppkomma på 1800-talet (Carlström & Nyman 2005). Dovvilt lever i ett brett spektrum av habitattyper, men blandlandskap av
jordbruksmark och löv- eller blandskogar verkar föredras. Det hindrar dock inte att dovvilt hittas i barrskogsdominerade områden och att barrskog kan utgöra en väsentlig andel av habitatsammansättningen i dovvilts hemområden i Sverige (Winsa 2008).
Arten klassas som en blandbetare (Hofmann 1989) där en stor del av födan kan utgöras av gräs, men också av örter, skott och blad liksom ek- och bokollon samt bark. Dovvilt söker sig gärna till åkermark där särskilt vete, havre, raps,
sockerbetor, potatis och vallodlingar utövar en hög attraktion (Chapman &
Chapman 1975; Putman m.fl. 1993; Carlström & Nyman 2005; Nichols m.fl. 2016; Rautiainen 2017).
Dovhindar kan i det vilda bli uppåt 20 år gamla. I regel föder de sin första kalv vid 2 års ålder. Hjortarna har en senare mognad, men också en betydligt kortare livslängd än hindarna och torde sällan bli äldre än cirka 10 år. Även om hjortarna är könsmogna vid 1,5 års ålder kan de egentligen inte betraktas som fullvuxna förrän de är cirka 4 år. Som starkast avseende konkurrenskraft under brunsten och hornstorlek är de i regel vid 6 - 9 års ålder.
Högbrunsten pågår under andra halvan av oktober och första halvan av november (Carlström & Nyman 2005; Lindström 2015). Under högbrunsten etablerar de dominanta hjortarna brunstplatser som förvaras mot andra hjortar. Till dessa relativt små brunstplatser (cirka 0,5 ha) försöker platshjortarna locka till sig hindar genom intensivt brölande. Dovviltet har därmed inget haremssystem, utan hindarna rör sig mellan olika platshjortar när de gör sitt val. Yngre hjortar och svagare hjortar rör sig mellan brunstplatser och i hindgrupper för att därigenom försöka få
kondition med följden att många av dessa dör efter brunsten eller under den påföljande vintern (Kjellander 2011).
Kalvarna föds i juni-juli (Kjellander m.fl. 2012). Dovhindar föder bara en kalv och dubbelkalvar är ovanligt. Under den första tiden ligger kalven gömd det mesta av tiden och det är först vid några veckors ålder som kalven börjar vistas i hind-kalvgrupper tillsammans med modern. Kalvarna kan samlas i grupper som övervakas av någon eller några hindar, samtidigt som resten av flocken söker bete någon annanstans.
Dovvilt är utpräglat grupplevande och kan bilda mycket täta populationer (Chapman & Chapman 1975; Putman m.fl. 1993; Putman 1996). Hindar och kalvar, ibland med inslag av yngre hjortar, bildar flockar som utgår från
grupperingar av närbesläktade hindar i olika generationer. Flockarna leds i regel av medelålders eller äldre hindar. Storleken på dessa hind-kalvflockar kan variera kraftigt beroende på populationstäthet och landskapsstruktur, men i öppna landskap med höga tätheter kan flockar på hundratals djur observeras. Utanför brunsttid lever hindar och hjortar segregerat. Hjortarna bildar särskilda så kallade
ungkarlsflockar, och söker sig ofta till områden där förekomst av hindar och kalvar är låg eller ingen (Chapman & Chapman 1975; Thirgood 1996; Carlström & Nyman 2005; Villerette m.fl. 2006).
Barriäreffekter dovvilt
Det är svårt att finna data på hur dovvilt påverkas av olika typer av barriärer. En delförklaring till detta är måhända den mycket långsamma spridningen till nya områden. I likhet med kronvilt kan utbredningen av dovvilt ha en gräns i en större väg (utan viltstängsel) och under flera år kan det finnas dovvilt på ena sidan vägen, men inte på den andra (Carlström och Nyman 2005). Dovvilt kan dock simma och vatten utgör alltså inte ett absolut hinder för spridning.
Studieområde dovvilt Sverige
Koberg (58°11´N, 12°41´E, boreonemoral zon) ligger i Västergötland cirka 12 km syd-sydost om Trollhättan. Terrängen är låglänt med 79 % skog (i huvudsak gran och tall), 16 % jordbruksmark och 2 % myr. Åkermarken används i stor
utsträckning för odling av vall för viltbete. En väg med viltstängsel delar in området i en nordlig och en sydlig del. Medeltätheten av dovvilt var under 2007 - 2015 för den nordliga delen 11,4/km2 och för den sydliga delen 30,8/km2
Rörelsemönster och hemområdesstorlek hos dovvilt
Dovviltets spridning till nya marker kan karaktäriseras som mycket långsam. Det verkar vanligt att hjortar lämnar födelseområdet under sitt andra eller tredje levnadsår, medan utvandringen av hindar verkar betydligt mer sparsam (Chapman & Chapman1975; Kjellander 2011). På Koberg utvandrade fyra av fem unga GPS-märkta hjortar vid en ålder av 1 - 3 år. Av sju hindar GPS-märkta som 1,5-åringar var det ingen som utvandrade. Materialet med kalvar som öronmärkts som nyfödda, kan dock eventuellt tyda på att det finns en utvandring av såväl hjortar som hindar före 1,5 års ålder (Kjellander 2011).
Sommarhemområden för hindar på Koberg är i genomsnitt 290 ha med 647 ha som störst uppmätta (Christensson 2010). För hjortarna på Koberg fann Lindström (2017) följande hemområdesstorlekar: vår 190 ha (40-840 ha), höst 770 ha (120 - 2330 ha), sommar 130 ha (30 - 560 ha), vinter 260 ha (60 - 850 ha), års inkl. brunst 650 ha (270 - 1 760 ha), års exkl. brunst 270 ha (140 - 660 ha). Av 18 hjortar gjorde fyra säsongsvandringar > 5 km mellan brunstplats och
vinter-sommarområde där den längsta vandringen var knappt 20 km (Lindström 2017).
Mellanartsinteraktioner älg, rådjur,
kronvilt och dovvilt
Även om älg, rådjur, kronvilt och dovvilt placeras olika längs skalan mellan ”browsers” (framförallt kvistätare) och ”grazers” (framförallt gräsätare), så kan födoöverlappet vara tämligen stort (för dovvilt - rådjur: Putman 1996; Putman m.fl.1996: Nichols m.fl. 2016, för dovvilt - kronvilt: Putman 1996; Putman m.fl.1996; Azorit m.fl. 2012, för kronvilt - rådjur: Matrai & Kabai 1989; Putman 1996; Putman m.fl.1996; Storms m.fl. 2008, för kronvilt - älg: Lavsund 1976; Mysterud 2000, för älg - rådjur: Mysterud 2000). I en sammanställning av
skandinaviska studier fann Mysterud (2000) ett dietöverlapp mellan älg och rådjur på 21 % sommartid och 34 % vintertid, medan dietöverlappet mellan älg och kronvilt vintertid var 32 %. På Koberg sågs ett dietöverlapp mellan dovvilt och rådjur på 79 % mätt som förekomst av olika födoslag. En kvantitativ analys visade ett överlapp på 30 % (Nichols m.fl. 2016).
Med ett stort överlapp vad gäller födoval, är det naturligt att de olika arterna också i stor utsträckning vistas i samma miljöer och kan ha ett betydande överlapp även avseende habitatutnyttjande (för dovvilt - rådjur: Putman 1996; Putman m.fl.1996; Ferretti m.fl. 2011a; Winsa 2008, för dovvilt - kronvilt: Putman 1996; Putman m.fl.1996; Ferretti & Mattiolo 2012; för kronvilt - rådjur: Putman 1996; Putman m.fl.1996; Prokesova m.fl. 2006; Borkowski & Ukalska 2008; Richard m.fl. 2013, för kronvilt - älg: Lavsund 1976). På fält med attraktiva grödor kan således kronvilt och dovvilt ses beta i varandras närhet, ibland till och med så flockarna blandas (Putman 1996; A. Jarnemo, pers. obs.). Likaså kan rådjur och älg ses beta ostört i absolut närhet till kronvilt (Lavsund 1976; A. Jarnemo, pers. obs.).
Alla fyra arter utnyttjar foderplatser. Ofta kan olika arter besöka samma foderplatser, och detta också samtidigt (Bartos m.fl. 1996; Ferretti m.fl. 2008, 2011a; Otto 2013; Johnsen 2015). Besöksfrekvens och tidpunkt för besök kan dock påverkas av förekomst av andra arter. Johnsen m.fl. (2015) såg att älg och kronvilt besökte samma foderplatser, ibland samtidigt, men visade också att ju mer en foderplats används av älg, desto mindre används den av kronvilt och vice versa. Otto (2013) fann att rådjur i större utsträckning besökte foderplatserna dagtid, jämfört med älg, kronvilt och dovvilt, och drog slutsatsen att rådjur har svårt att komma till i närvaro av de andra arterna, eller att de aktivt körs undan. På Koberg fann Cederholm (2012) samma mönster att rådjur utnyttjade foderplatser in mindre utsträckning i närvaro av dovvilt. Framförallt dovvilt visar aggressivt beteende och har konstaterats tränga undan såväl rådjur som kronvilt vid foderplatser (Bartos m.fl. 1996; Ferretti 2011; Ferretti m.fl. 2008, 2011a). På Koberg har det också observerats att med ökande gruppstorlek hos dovvilt, så höll sig rådjuren på längre avstånd från dovviltet (Agnrud 2011). Under kronviltets brunst kan man dock se att platshjortar kan köra iväg dovhjortar från sin närhet (Jarnemo 2017).
Tätheten av rådjur har visat sig negativt korrelerad till täthet av dovvilt, med de högsta rådjurstätheterna där tätheten av dovvilt är låg (Agnerud 2011; Ferretti m.fl. 2011a). Konkurrens från dovvilt har visat sig ha en negativ påverkan på
habitatkvalitet för rådjur (Focardi m.fl. 2006) och en negativ effekt av dovvilt på rådjur härrör troligen både från födokonkurrens och från interferens (Focardi m.fl. 2006; Ferretti m.fl. 2011a, 2011b; Kjellander 2011). Även kronvilt verkar påverkas negativt av konkurrens med dovvilt (Azorit m.fl. 2012; Ferretti & Mattiolo 2012). Rådjur kan under vissa omständigheter också påverkas negativt av kronvilt. En ökad täthet av kronvilt i ett område med näringsfattig vegetation, ledde till lägre vikter hos såväl vuxna rådjur som kid (Richard m.fl. 2010). Andra studier har dock inte kunnat påvisa någon konkurrens mellan kronvilt och rådjur, trots betydande habitat- och dietöverlapp (Prokesova m.fl. 2006; Borkowski & Ukalska 2008; Storms m.fl. 2008; Richard m.fl. 2013).
Källförteckning
Agnrud A. 2011. Konkurrens mellan rådjur och dovhjort. - Examensarbete, SLU-Skogsmästarskolan, Skinnskatteberg.
Allen AM. 2016. Linking movement ecology with the management of mobile species. - Doktorsavhandling, Sveriges lantbruksuniversitet,
Allen AM, Månsson J, Sand H, Malmsten J, Ericsson G, Singh NJ. 2016. Scaling up movements: From individual space use to population patterns. -
Ecosphere 7:e01524. DOI: 10.1002/ecs2.1524.
Azorit C, Tellado S, Oya A, Moro J. 2012. Seasonal and specific diet variations in sympatric red and fallow deer of southern Spain: a preliminary approach to feeding behavior. - Animal Production Science 52: 720-727.
Bartos L, Vankova D, Siler J, Losos S. 1996. Fallow deer tactic to compete over food with red deer. - Aggressive Behavior 22: 375-385.
Bartzke GS, May R, Solberg EJ, Rolandsen CM, Roskaft E. 2015. Differential barrier and corridor effects of power lines, roads and rivers on moose (Alces
alces) movements, - Ecosphere 6: 67.
Bateson P, Bradshaw EL. 1997. Physiological effects of hunting red deer (Cervus
elaphus). - Proceedings of the Royal Society London B. 264: 1707-1714.
Bevanda M, Fronhoffer EA, Heurich M, Müller J, Reineking B. 2015. Landscape configuration is a major determinant of home range size variation. - Ecosphere 6: 195.
Bonenfant C, Loe LE, Mysterud A, Langvatn R, Stenseth NC, Gaillard, J-M, Klein, F. 2004. Multiple causes of sexual segregation in European red deer:
enlightenments from varying breeding phenology at high and low latitude. - Proceedings of the Royal Society London B 271: 883-892.
Borkowski J, Ukalska J. 2008. Winter habitat use by red and roe deer in pine-dominated forest. - Forest ecology and management 255: 468-475.
Bramley PS. 1970. Territoriality and reproductive behaviour of roe deer. - Journal of Reproduction and Fertility 11: 43-70.
Bresinski W. 1982. Grouping tendencies in roe deer under agrocenosis conditions. - Acta Theriologica 27: 427-447.
Carlström L, Nyman M. 2005. Dovhjort. Jägareförlaget/Svenska Jägareförlaget. Catchpole EA, Fan Y, Morgan BJT, Clutton-Brock TH, Coulson T. 2004. Sexual
dimorphism, survival and dispersal in red deer. - Journal of Agricultural, Biological and Environmental Statistics 9: 1-26.
Cederholm, T. 2012. Use and competition at artificial feeding sites – the roe deer and fallow deer case. - BSc-thesis Department of Ecology, SLU. 2012:11. Cederlund G, Liberg O. 1995. Rådjuret. Viltet, ekologin och jakten. Svenska
Jägareförbundet.
Chapman D, Chapman N. 1975. Fallow deer. Coch-Y-Bonddu Books, Machynlleth Clutton-Brock TH, Albon SD, Guinness FE. 1988. Reproductive success in male
and female red deer. - In: Reproductive success (ed.T.H. Clutton-Brock), s. 325–343. Chicago, IL: The University of Chicago Press.
Clutton-Brock TH, Guinness FE, Albon SD. 1982. Red deer – behaviour and ecology of two sexes. Edinburgh University Press, Edinburgh.
Clutton-Brock TH, Iason GR, Guinness FE. 1987. Sexual segregation and density-related changes in habitat use in male and female red deer (Cervus elaphus). - Journal of Zoology (Lond) 211: 275-289.
Coulon A, Cosson F, Angibault M, Cargnelutti B, Galan, M, Morelett, N, Petit, E, Aulagnier S, Hewison AJM. 2004. Landscape connectivity influences gene flow in a roe deer population inhabiting a fragmented landscape: an individual-based approach. - Molecular Ecology 13: 2841-2850. Coulon A, Guillot G, Cosson F, Angibault JMA, Aulagnier, S, Cargnelutti B,
Glaan M, Hewison AJM. 2006. Genetic structure is influenced by landscape features: empricial evidence from a roe deer population. - Molecular
Ecology 15: 1669-1679.
Christensson M. 2010. Despotic distribution in female fallow deer (Dama dama) groups. - Report Independent project biology, SLU, Grimsö.
Daniels M, McClean C. 2002. Red deer calf tagging programmes in Scotland – an analysis. - Deer 12: 420-423.
Danilkin A, Hewison AJM. 1996. Behavioural Ecology of Siberian and European roe deer. Chapman and Hall, London.
Davis M, Stephens P, Kjellander P. 2016. Beyond Climate Envelope Projections: Roe Deer Survival and Environmental Change. - Journal of Wildlife Management 80: 452-464.
Debeffe L, Focardi S, Bonenfant C, Hewison AJM, Morellet N, Vanpé C, Heurich M, Kjellander P, Linnell JDC, Mysterud A, Pellerin M, Sustr P, Urbano F, Cagnacci F. 2014. A one night stand? Reproductive excursions of female roe deer as a breeding tactic. - Oecologia 176: 431-443.
Duncan P, Tixier H, Hoffman RR, Lechner-Doll M. 1998. Feeding strategies and the physiology of digestion in roe deer. - In: Andersen R, Duncan P, Linnell JDC (eds.), The European roe deer: The biology of success. Scandinavian University Press, Oslo, Norway, pp. 91-116.
Ericsson G, Wallin K. 2001. Age-specific moose Alces alces mortality in a
predator free environment: evidence for senescence in females. - Écoscience 8:157–163.
Ericsson G, Boman M, Mattson L. 2000. Selective versus random moose
harvesting: does it pay to be a prudent predator? - Journal of Bioeconomics 2:117-132.
Ericsson G, Wallin K, Ball JP, Broberg M. 2001. Age-related reproductive effort and senescene in free-ranging moose Alces alces. - Ecology 82:1613-1620. Ericsson G, Neumann W, Stenbacka F, Evans A, Pettersson J, Dettki H, Arnemo
JM, Singh N, Jatko M, Sundgren B, Saitzkoff R. 2017b. Årsrapport GPS-älgarna Norrbotten 2016-2017; vandring och rörelseaktivitet i
Haparanda-Ericsson G, Neumann W, Stenbacka F, Evans A, Pettersson J, Dettki H, Arnemo JM, Cromsigt J, Singh N. 2016b. Slutrapport GPS-älgarna i Tjåmotis 2014-2016; rörelse och reproduktion. Sveriges Lantbruksuniversitet, 27pp. Ericsson G, Neumann W, Allen A, Stenbacka F, Evans A, Pettersson J, Dettki H,
Arnemo JM, Cromsigt J, Singh N, Jatko M, Sundgren B, Saitzkoff R, Kindberg J. 2016a. Slutrapport GPS-älgarna Norrbotten 2013-2015; rörelse och reproduktion i Arvidsjaur, Niemisel, och Ängesån. Sveriges
lantbruksuniversitet, 29pp
Espmark Y. 1969. Mother-young relations and development of behaviour in roe deer (Capreolus capreolus L.). - Viltrevy 6: 461-540.
Ferretti F. 2011. Interspecific aggression between fallow and roe deer. - Ethology Ecology & Evolution 23: 179-186.
Ferretti F, Mattioli S. 2012. The Mesola red deer: present numbers and conservation perspectives. - Hystrix 23: 35-43.
Ferretti F, Sforzi A, Lovari S. 2008. Intolerance amongst deer species at feeding: Roe deer are uneasy banqueters. - Behavioural Processes 78: 487-491. Ferretti F, Bertoldi G, Sforzi A, Fattorini L. 2011a. Roe and fallow deer: are they
compatible neighbours? - European Journal of Wildlife Research 57: 775-786.
Ferretti F, Sforzi A, Lovari S. 2011b. Behavioural interference between ungulate species: roe are not on velvet with fallow deer. - Behavioral Ecology and Sociobiology 65: 875-887.
Ferretti F, Sforzi A, Lovari S. 2012. Avoidance of fallow deer by roe deer may not be habitat-dependent. - Hystrix 23: 28-34.
Focardi S, Aragno P, Montanaro P, Riga F. 2006. Inter-specific competition from fallow deer Dama dama reduces habitat quality for the Italian roe deer
Capreolus capreolus italicus. - Ecography 29: 407–417.
Frantz AC, Bertouille S, Eloy MC, Licoppe A, Chaumont F, Flamand MC. 2012. Comparative landscape genetic analyses show a Belgian motorway to be a gene flow barrier for red deer (Cervus elaphus), but not wild boars (Sus
scrofa). - Molecular Ecology 21: 3445-3457.
Frantz AC, Hamann, JL, Klein F. 2008. Fine-scale genetic structure of red deer (Cervus elaphus) in a French temperate forest. - European Journal of Wildlife Research 54: 44-52.
Gaillard JM, Hewison AJM, Kjellander P, Pettorelli N, Bonenfant C, Van Mooster B, Liberg O, Andren H, Van Laere G, Klein F, Angibault JM, Coulon A, Vanpé C. 2008. Population density and sex do not influence fine-scale natal dispersal in roe deer. - Proceedings of the Royal Society B. Biological Sciences, 275: 2025-2030.
Gebert C, Verheyden-Tixier H. 2001. Variations of diet composition of red deer (Cervus elaphus L.) in Europe. - Mammal Review 31: 189-201.
Georgii B, Peters-Ostenberg E, Henneberg M, Herrmann M, Müller-Stieß H, Bach L. 2007. Nutzung von Grünbrücken und anderen Querungsbauwerken durch Säugetiere. Gesamtbericht zum Forschungs-und Entwicklungsvorhabe. Forschung Strassenbau und Strassenverkehrstechnik Heft 971. Bonn.
Guillet C, Bergström R, Cederlund G. 1996. Size of winter home range of roe deer
Capreolus capreolus in two forest areas with artificial feeding in Sweden. -
Wildlife Biology 2: 107-111.
Haanes, H, Røed, KH, Rosef, O. 2011. Sex-biased dispersal in a northern ungulate population. - Rangifer 31: 139-153.
Hewison AJM, Vincent JP, Reby D. 1998. Social organization of European roe deer. - In: Andersen R, Duncan P, Linnell JDC (eds.), The European roe deer: The biology of success. Scandinavian University Press, Oslo, Norway, pp. 189-220.
Hofmann RR. 1989. Evolutionary steps of ecophysiological adaptation and diversification of ruminants: comparative view of their digestive system. - Oecologia, 78: 443–457.
Höglund J, Cortazar-Chinarro M, Jarnemo A, Thulin C-G. 2013. Genetic variation and structure in Scandinavian red deer (Cervus elaphus): influence of ancestry, past hunting, and restoration management. - Biological Journal of the Linnean Society 109: 43-53.
Jarnemo A. 2008. Seasonal migration of red deer males in southern Sweden and consequences for management. - European Journal of Wildlife Research 54: 327-333.
Jarnemo A. 2011. Male red deer (Cervus elaphus) dispersal during the breeding season. - Journal of Ethology 29: 329-336.
Jarnemo A. 2014. Kronviltprojektet 2005-2013. Slutrapport. Grimsö forskningsstation, SLU.
Jarnemo A. 2017. Ögonblicket: Kronhjorten och dovspetsarna. Svensk Jakt 10: 22-23.
Jarnemo A, Wikenros C. 2014. Movement pattern of red deer during drive hunts in Sweden. - European Journal of Wildlife Research 60: 77-84.
Jeppesen JL. 1987. Impact of human disturbance on home range, movements and activity of red deer (Cervus elaphus) in a Danish environment. - Danish Review of Game Biology 13: 1-38.
Johansson A. 1996. Territory dynamics and marking behaviour in male roe deer. - Ph.D. thesis, Stockholm University.
Johnsen K, Zimmerman B, Milner J, Wedul S. 2015. Vil hjorten utkonkurrere elgen på fôringsstasjonene? WWW.HJORTEVILT.NO, nedladdad 2018-01-16.
Kindbladh N. 2015. Timing of the rut in fallow deer Dama dama. - Master thesis, SLU, Grimsö.
Kjellander P. 2000. Density dependence in roe deer population dynamics. - Doctoral thesis, Swedish University of Agricultural Sciences, Uppsala. Kjellander P. 2011. Lägesrapport. Konkurrens mellan stora växtätare: exemplet