Malmö högskola
Lärarutbildningen
Skolutveckling och ledarskap
Examensarbete
10 poängLyssnar läroboksförfattaren på forskaren?
Do Textbook Authors Listen to Scientists?
Johan Wahlin
Lärarexamen 60 poäng Lärarutbildning 60p 2007-10-16
Examinator: Lena Lang
Wahlin, Johan. (2007). Lyssnar läroboksförfattaren på forskaren? (Do Textbook Authors Listen to Scientists?). Natur, Miljö, Samhälle. Lärarutbildningen, Malmö högskola.
Sammanfattning
En rad områden som elever finner särskilt svåra har identifierats inom naturvetenskaplig ämnesdidaktisk forskning de senaste decennierna. Syftet med den här studien är att undersöka i vilken mån naturvetenskapliga läroböcker är skrivna på ett sätt som kan tänkas överbrygga dessa kända svårigheter. Dessutom undersöktes om det i detta avseende har skett några förändringar i de läroböcker som publicerats efter forskningsresultaten blev kända jämfört med dem som publicerats tidigare. Genetikavsnitten i två nypublicerade och en äldre biologilärobok analyserades med avseende på huruvida kända svårigheter överbryggas eller inte. Analysen visade att lärobokförfattarna inte på ett nämnvärt sätt undvikit svårigheterna. De nyare böckerna var inte heller bättre i detta avseende än den äldre boken.
Nyckelord: genetik, biologi, vardagsföreställningar, läromedelsanalys, förförståelse, svårigheter, naturvetenskap, läroböcker
Innehållsförteckning
1. Inledning ……….……. 7
2. Syfte och frågeställningar ……….…….. 8
3. Teoretisk bakgrund ……..……….. 9
3.1 Eleven i fokus ……… 9
3.2 Didaktiska problem inom naturvetenskapsundervisning ……… 10
3.3 Didaktiska problem inom genetikundervisning ………...… 12
3.3.1 Ämnet innehåller många fackuttryck ………...……. 13
3.3.2 Matematiska beräkningar inom ämnet kan vara komplicerade …...………. 14
3.3.3 Cellulära processer kan vara svåra att förstå ………...……. 14
3.3.4 Ämnet kan bli abstrakt beroende på undervisningens upplägg ……...……. 15
3.3.5 Snabba hopp mellan makro- och mikronivå kan vara förvirrande …...……. 16
3.4 Vad måste man kunna för att förstå genetik? ………..… 16
3.5 Hur har läroböcker förändrats över tiden? …………...………..… 17
4. Material och metod ……..………... 19
4.1 Val av metod……...………..……….………….… 19
4.2 Avgränsningar och urval ……….………….… 20
4.3 Om böckerna ……….……… 22
4.3.1 Spira –Biologi A ………....……. 22
4.3.2 Biologi A med naturkunskap ………...……. 22
4.3.3 Biologi för gymnasieskolan ……….……...….…. 23
4.4 Metod ……….……… 24
4.4.1 Ämnet innehåller många fackuttryck ………...……. 24
4.4.2 Matematiska beräkningar inom ämnet kan vara komplicerade …...………. 25
4.4.3 Cellulära processer kan vara svåra att förstå ………...…..…. 25
4.4.4 Ämnet kan bli abstrakt beroende på undervisningens upplägg ……..….…. 25
4.4.5 Snabba hopp mellan makro- och mikronivå kan vara förvirrande ….…..…. 25
5. Resultat ……..……….. 27
5.1 Ämnet innehåller många fackuttryck……….….…. 27
5.2 Matematiska beräkningar inom ämnet kan vara komplicerade………. 31
5.3 Cellulära processer kan vara svåra att förstå……...…………...…….... 32
5.4 Ämnet kan bli abstrakt beroende på undervisningens upplägg……... 32
5.5 Snabba hopp mellan makro- och mikronivå kan vara förvirrande….... 33
5.6 Sammanfattning av resultaten………... 33
6. Diskussion ……..……….. 36
6.1 Ämnet innehåller många fackuttryck……….….…. 37
6.2 Matematiska beräkningar inom ämnet kan vara komplicerade………. 40
6.3 Cellulära processer kan vara svåra att förstå……...…………...…...…. 41
6.4 Ämnet kan bli abstrakt beroende på undervisningens upplägg……….. 42
6.5 Snabba hopp mellan makro- och mikronivå kan vara förvirrande….... 43
7. Avslutning ……..………...……….….. 45
7.1 Lyssnar läroboksförfattaren på forskaren?... 45
7.2 Frågor för framtiden... 46
Referenser ……..………..………..….. 47
Bilagor ……..……….….. 51
1. Inledning
De senaste decennierna har en lång rad ämnesdidaktiska studier publicerats som undersökt hur elever tänker om olika naturvetenskapliga fenomen (se t.ex. Projekt Nordlab). Många områden som elever finner extra svåra har kunnat identifieras och sätt att överbrygga dessa svårigheter har föreslagits. En förutsättning för att dessa förslag kommer till användning är naturligtvis att forskningsresultaten når ut i klassrummen. Om dessa resultat fanns inkorporerade i läroböcker skulle sannolikt en del av de problem som har identifieras kunna undvikas bland svenska elever. Undersökningar har nämligen visat att en stor majoritet av de svenska lärarna regelbundet använder sig av läroböcker, medan under två procent av lärarna säger sig aldrig använda dem (Levén, 2003).
Eftersom jag själv har undervisat både universitetsstudenter och gymnasieelever i genetik har jag en speciell förkärlek för detta ämne. Genetikämnet är också tillsammans med ett annat ämne den del av biologin som elever upplever som svårast (Bahar et al., 1999). Mot bakgrund av detta och att undersökningen annars skulle bli alldeles för stor har jag valt att begränsa det här arbetet till att omfatta gymnasieskolans genetikundervisning. Jag har valt att analysera ett par nyare biologiböcker för att undersöka hur de kända svårigheterna hanteras. Dessutom har jag valt att jämföra dessa resultat med hur svårigheterna hanterades i böcker som publicerades innan forskningsresultaten blev kända, för att se om det har skett någon förbättring över tiden i detta avseende.
Jag tror att analyser av det här slaget är nödvändiga för att ta reda på i vilken mån den omfattande ämnesdidaktiska forskning som bedrivits de senaste decennierna kommer eleverna till godo. Det är ju trots allt för deras skull som undervisningen bedrivs.
2. Syfte och frågeställningar
Den naturvetenskapliga ämnesdidaktiska forskningen har under de senaste årtiondena identifierat många områden som elever finner särskilt komplicerade och föreslagit metoder för att undvika dessa problem i undervisningen.
Syftet med den här studien är att undersöka i vilken mån naturvetenskapliga läroböcker är skrivna på ett sätt som kan tänkas överbrygga dessa kända svårigheter. Dessutom undersöks om det i detta avseende har skett några förändringar i de läroböcker som publicerats efter forskningsresultaten blev kända jämfört med dem som publicerats tidigare.
De frågor som den här studien försöker besvara är sålunda:
• Hanterar läroböcker de svårigheter som identifierats i den ämnesdidaktiska forskningen på ett tillfredsställande sätt?
• Har det i detta avseende skett någon förändring över tiden?
Undersökningen omfattar litteraturstudier av genetikavsnitten i tre läroböcker avsedda för gymnasieskolans Biologi A-kurs. Mer om denna och andra avgränsningar beskrivs senare.
3. Teoretisk bakgrund
3.1 Eleven i fokus
Innan de ämnesdidaktiska problemen presenteras kan det vara på sin plats att diskutera den socialkonstruktivistiska kunskapssynen och hur den har kommit att sätta eleven i fokus. Socialkonstruktivismen kombinerar Jean Piagets och Lev Vygotskys teorier om hur kunskap blir till (Andersson, 2001; Sjøberg, 2005).
Denna kunskapssyn har sin grund i konstruktivismen, en term som brukar tillskrivas Jean Piaget (Andersson, 2001; Sjøberg, 2005). Enligt denna kunskapssyn sätter all mental aktivitet igång ett antal processer som konstruerar minnen, föreställningar och begrepp.
När en person möter något fenomen som inte stämmer överens med personens uppfattning om fenomenet, rubbas en intern jämvikt. Denna rubbade jämvikt kommer att återställas genom att personen modifierar sin uppfattning om fenomenet och därmed återställer jämvikten. Denna process kallas jämvikt genom självreglering och är en viktig del i Piagets konstruktivism. I den naturvetenskapliga undervisningen kan läraren använda denna jämvikt genom självreglering genom att utmana eleven att t.ex. försöka förutse resultatet av en laboration (Andersson, 2001). Läraren kan således genom att utmana elevens föreställningar hjälpa denna att modifiera sina kunskaper inom ett visst ämne.
En annan viktig komponent i Piagets konstruktivism är att människan till naturen är nyfiken och vetgirig (Andersson, 2001). Detta innebär ur ett lärarperspektiv att eleven
vill lära sig nya saker, det gäller bara att hitta ett sätt att stimulera honom eller henne.
En tredje viktig komponent i Piagets teorier är tanken att hjärnan skapar strukturer. Minnen, föreställningar och begrepp konstrueras när dessa strukturer är aktiverade. Piaget menade också att dessa strukturer skiljer sig från varandra under olika delar i en persons liv. Det kan alltså vara fördelaktigt att som lärare förstå att yngre barn, och tonåringar kommer att tänka på ett delvis annat sätt än en vuxen person. Med Piaget
kom eleven att sättas i fokus i skoldebatten. Det handlade nu om att hjälpa eleven att konstruera sin egen kunskap snarare än att försöka överföra kunskap från lärare till elev.
På 1980-talet uppmärksammades Lev Vygotskys teorier kring kunskap (Andersson, 2001). Vygotsky menade att begrepp som t.ex. gen eller kromosom är socialt medierade. En person kan inte själv upptäcka dessa termer, utan för att nå kunskap om dem krävs en social dimension. Personen måste träffa på människor som använder och diskuterar termerna, för att på så sätt själv förstå vad de innebär.
Inom socialkonstruktivistisk kunskapsteori, som alltså är en kombination av Piagets och Vygotskys teorier, blir kunskap individuellt konstruerad och socialt medierad (Andersson, 2001).
I och med socialkonstruktivismens framväxt började ämnesdidaktiker fråga sig hur elever tänker om olika naturvetenskapliga fenomen (Andersson, 2001). Man började också skilja på elevers vardagsföreställningar om fenomenen och de förklaringar som naturvetenskaplig forskning presenterade och undersöka hur dessa skiljer sig från varandra. Dessutom började man presentera didaktiska metoder för att hantera de områden som elever ofta upplever som problematiska.
3.2 Didaktiska problem inom naturvetenskapsundervisning
Socialkonstruktivismens framväxt ledde till att eleven hamnade i centrum och att man inom den ämnesdidaktiska forskningen började intressera sig för hur den enskilde eleven tänker om olika fenomen samt vilka områden som han eller hon finner särskilt svåra.
Eftersom de naturvetenskapliga ämnena ofta upplevs som särskilt komplicerade och abstrakta är det inte förvånande att mycket av den ämnesdidaktiska forskningen ägnas åt att beskriva problem som elever och lärare upplever just inom detta ämnesområde. Sanders (1993) delar upp dessa problem i tre olika kategorier:
2. Problem som beror på att eleven missförstår kursstoffet
3. Problem som uppstår av felaktig information från lärare eller läromedel.
I skrivande stund innehåller Reinders Duits bibliografiska databas referenser till ca 7700 vetenskapliga artiklar som behandlar just detta ämne (Duit, 2004). En stor del av dessa artiklar behandlar elevers vardagsföreställningar av naturvetenskapliga fenomen (Wallin, 2004). Det ligger sannolikt i människans natur att från enkla observationer bilda lagar för hur saker och ting fungerar. Sådana lagar hjälper oss nog ofta att klara oss i en komplicerad värld, men kan också leda fel. En vuxen person kan titta ut genom fönstret och konstatera att vinden får trädkronorna att röra sig. Samma observation kan hos ett barn tolkas som att trädkronorna ruskar sig så att ett vinddrag uppstår (Andersson, 2003a).
Ett annat exempel på hur vardagsföreställningar kan leda till felaktiga slutsatser framkommer när elever tillfrågas om vad som händer då man höjer temperaturen på en platta på vilken det står en kastrull med kokande vatten (Andersson, 2003a). Elever kan då svara att temperaturen på vattnet i kastrullen kommer att höjas. Detta kan tyckas vara en rimlig slutsats: ju mer värme från plattan, desto varmare vatten. Likväl leder också detta resonemang fel.
Att eleven missförstår kursstoffet har inte som vardagsföreställningarna någon positiv funktion. Därför är det upp till läromedelsförfattare och lärare att göra allt som står i deras makt för att förhindra att sådana missförstånd uppstår. Många forskningsstudier fokuserar just på vad elever ofta har problem med inom de naturvetenskapliga ämnesområdena. När 700 niondeklassare fick förklara vad det beror på att månen ibland är hel och ibland är halv framkom flera olika förklaringsmodeller (Andersson, 2003b). Fyra ungefär lika vanligt förekommande modeller stod för 73 % av svaren:
1. Något (moln, sol eller planeter) täcker för månen. 2. Jorden blockerar ljuset från solen.
3. Beror på hur solen skiner på månen
4. Solen lyser på halva månen. Hur vi uppfattar månen beror på vinkeln vi betraktar månen ifrån.
Det torde vara uppenbart att man skulle ha stor nytta av att känna till dessa vanliga missförstånd innan man som lärare inleder undervisningen kring vårt solsystem. Genom att känna till dem kan undervisningen läggas upp på ett sätt som gör att eleverna kan undvika de vanligaste fallgroparna. Liknande exempel är vanliga inom naturvetenskaplig ämnesdidaktisk forskning. Inom ramarna för projektet Nordlab har en rad sådana exempel tagits fram (Projektet Nordlab). Detta projekt, som genomdrevs mellan åren 2000 och 2003, var ett nordiskt samarbetsprojekt vars syfte var att ge lärare inom framförallt de naturvetenskapliga ämnena redskap för att förbättra sin undervisning. Projektet Nordlab resulterade bl.a. i ett antal workshops med övningar som finns tillgängliga för nedladdning från hemsidan.
Den tredje av Sanders problemkategorier, identifierar problem som uppstår genom felaktiga uppgifter från lärare eller läromedel. Det är troligtvis en omöjlig uppgift att helt utradera alla felaktiga uppgifter som elever utsätts för. Icke desto mindre kan man vidta steg för att minska dessa. Läraren kan inte veta allting om sitt ämne och kan nog ibland ha svårt att veta sina begränsningar. Genom kontinuerlig kompetensutveckling kan sannolikt en del av dessa missförstånd elimineras. För läromedelsförfattarna är det viktigt att vara noggranna och att få sitt material granskat av utomstående experter. I undervisningen kan man använda sig av flera olika läromedel, vilket gör det lättare att identifiera felaktiga uppgifter eftersom de olika källorna sannolikt kommer att helt eller delvis motsäga varandra.
3.3 Didaktiska problem inom genetikundervisning
Eftersom undersökningen i den här studien kommer att behandla analyser av genetikavsnitt ur läroböcker följer här en beskrivning av de problem som framkommit inom undervisningen av detta ämne.
Knippels (2002) har sammanfattat de didaktiska problemen inom genetikundervisning i fem punkter:
1. Ämnet innehåller många fackuttryck.
3. Cellulära processer kan vara svåra att förstå.
4. Ämnet kan bli abstrakt beroende på undervisningens upplägg. 5. Snabba hopp mellan makro- och mikronivå kan vara förvirrande.
3.3.1 Ämnet innehåller många fackuttryck
Facktermerna inom genetikämnet är många och komplexa. Dessutom liknar många av orden varandra trots att de har olika betydelse (Bahar et al., 1999). Ett par sådana exempel är kromosom-kromomer-kromatid och
nukleotid-nukleosid-nukleol-nukleinsyra. Fackuttrycken blir extra många på grund av att många av termerna har
synonymer som används parallellt. Exempel på sådana är gen och arvsanlag samt allel och genvariant. Problemet med de många, och komplexa fackuttrycken kan göra det svårt för eleverna att tillgodogöra sig genetikämnet och leda till att de missförstår kursstoffet. Sådana missförstånd utgör en av de kategorier som Sanders (2003) har identifierat som problemområden inom naturvetenskaplig undervisning (se ovan). Det är inte bara eleverna som upplever de många termerna som förvirrande. Även lärare kan uppleva dem som besvärliga (Andersson, 2003c). Dessutom är det vanligt att lärare, även på universitetsnivå, slarvar med termerna och därmed använder dem på ett felaktigt sätt, vilket leder in på Sanders tredje problemområde: de som är resultatet av felaktig information från lärare eller läromedel. Ett sådant exempel är när termerna gen och allel används synonymt (Cho et al., 1985). Med allel menas genvariant. Människan har t.ex. en gen som styr ögonfärgen. Det finns två alleler för ögonfärg: en som ger blåa ögon och en som ger bruna. Det är dock vanligt att lärare eller läromedel felaktigt hävdar att det finns två gener för ögonfärgen hos människan.
En del av fackuttrycken kan missförstås av elever på grund av att termerna har andra betydelser utanför vetenskapen. Ett sådant exempel är uttrycket dominant som används om alleler och hos en del elever får representera en god, stark egenskap medan en
recessiv allel får en negativ innebörd (Pearson & Hughes, 1988). Detta missförstånd
beror sannolikt på att eleverna använder sig av sina vardagsföreställningar (se Sanders första problemkategori ovan).
3.3.2 Matematiska beräkningar inom ämnet kan vara komplicerade
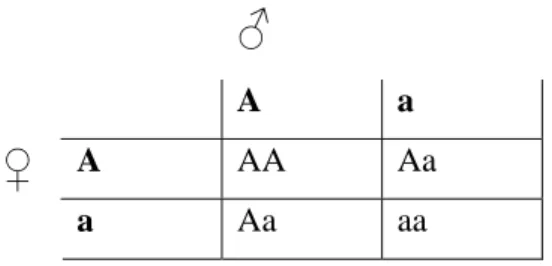
De uträkningar som används inom skolgenetiken är främst av sannolikhetskaraktär. Dessa uppgifter ställer inte bara till problem för elever som har svårt för matematik utan även för en del elever som hanterar sannolikheter på ett korrekt sätt i andra sammanhang (Kinnear, 1983). En orsak till detta tycks vara sättet på vilket symboler används vid uppgifterna (Bahar et al., 1999). Symbolerna används ibland på ett icke-intuitivt och inkonsekvent sätt. Inom den ämnesdidaktiska forskningen har man bl.a. lyft fram s.k. korsningsscheman (se fig. 3.1) som särskilda problem (Kinnear, 1983). Korsningsscheman används för att räkna ut vad sannolikheten är för att avkomman kommer att få en viss allelkombination från sina föräldrar.
♂
A a
A AA Aa
♀
a Aa aa
Figur 3.1. Korsningsschema. Normalt används
bokstavskombinationerna som ovan: stor bokstav representerar en dominant allel medan liten bokstav representerar en recessiv allel.
Elever begår ofta misstaget att tolka korsningsdiagram deterministiskt (Collins & Stewart, 1989). Låt oss använda oss av ett exempel på detta. Vi antar att en brunögd kvinna med en allel för brunögdhet och en allel för blåögdhet får barn med en brunögd man med likadan alleluppsättning. Barnet kommer då att ha 75 % sannolikhet att få bruna ögon och 25 % sannolikhet att få blåa ögon. Om paret har tre brunögda barn skulle en person som tolkar denna information deterministiskt säga att det fjärde barnet kommer att få blåa ögon. Detta överensstämmer naturligtvis inte med verkligheten eftersom sannolikheten att det fjärde barnet kommer att få blåa ögon fortfarande bara är 25 %.
3.3.3 Cellulära processer kan vara svåra att förstå
Flera studier har kommit fram till att den del av den klassiska genetiken som elever upplever som svårast är att förstå hur celldelningen fungerar (Bahar et al., 1999; Lewis
et al., 2000). Det finns två huvudsakliga sorters celldelning: mitos och meios. Med
mitos menas den celldelning som sker överallt i kroppen och resulterar i en identisk dottercell. Meiosen sker i testiklar och äggstockar och är en del i bildandet av könsceller (ägg och spermier). Elever upplever särskilt den senare processen som svårbegriplig, vilket tycks bero på en undermålig förståelse av kromosombegreppet (Lewis et al., 2000). Problemet förvärras av att elever ofta blandar ihop kromosom- och kromatidbegreppen. Med anledning av detta rekommenderar Longdon (1982) att begreppet kromatid undviks eftersom det inte bidrar till förståelsen av det biologiska fenomenet.
När Cho et al. (1985) undersökte tre olika amerikanska biologiläroböcker fann de att meiosen och den klassiska genetiken behandlas i olika delar av böckerna. Dessutom kopplades inte dessa nära besläktade ämneskategorier ihop, vilket sannolikt försvårar elevernas förståelse av fenomenen. Knippels et al. (2005) rekommenderar att man redovisar separata cellcykler för somatiska cellinjer (mitos) och könscellslinjer (meios).
Bahar et al. (1999) menar att man inte bör lägga upp undervisningen så att man avhandlar mitos och meios direkt efter varandra eftersom detta späder på elevernas uppfattning att dessa fenomen är nästan identiska.
3.3.4 Ämnet kan bli abstrakt beroende på undervisningens upplägg
Genetikämnet kan uppfattas som abstrakt av elever om de olika delmomenten behandlas i fel ordning i undervisningen. Ett sådant problem mötte vi ovan (3.3.3) när meiosen och den klassiska genetiken behandlades separat från varandra (Cho et al., 1985).
Symbolspråket inom genetikämnet kräver att eleverna besitter ett välutvecklat abstrakt tänkande. Som vi såg ovan (3.3.2) används ofta inte symbolerna för alleler på ett konsekvent sätt (Kinnear, 1983). Även symboler som används vid släktträd (pedigrees) och korsningsscheman har identifierats som problematiska för eleverna.
3.3.5 Snabba hopp mellan makro- och mikronivå kan vara
förvirrande
Ämnet genetik innehåller många storleksnivåer. En gen verkar på molekylär nivå, men påverkar individen på makronivå. Sådana hopp kan vara komplicerade för elever att följa. Dessutom förklaras fenomen ofta bara på individnivå, och inte på molekylär nivå (Bahar et al., 1999). Marbach-Ad & Stavy (2000) och Knippels et al. (2005) rekommenderar att man börjar undervisningen på makronivå för att sedan gå ner på lägre nivåer. Bahar et al. (1999) menar att man bör använda en storleksnivå i taget och inte hoppa mellan makro- och mikronivå, medan Knippels et al. (2005) menar att det viktiga är att man använder hopp mellan nivåer på ett konsekvent och logiskt sätt.
3.4 Vad måste man kunna för att förstå genetik?
En förutsättning för att bedriva framgångsrik undervisning i genetik är naturligtvis att man som lärare har klart för sig vad man vill att eleverna ska kunna. Inom skolan är det främst kursplanerna som definierar dessa mål. Genetikundervisningen bedrivs till största delen inom kursen Biologi A för gymnasieskolan. I kursplanen för Biologi A lyder målen som direkt relaterar till genetik (Skolverket, 2000):
”Eleven skall […] ha kunskap om arvsmassans strukturer samt förstå sambanden mellan dessa och individens egenskaper”
”Eleven skall […] ha kunskap om gentekniska metoder och deras tillämpningar samt kunna diskutera genteknikens möjligheter och risker ur ett etiskt perspektiv.”
Motsvarande mål för elever som gått ut det nionde skolåret lyder (Skolverket, 2000):
”Eleven skall […] ha kännedom om det genetiska arvet”
Inom ramen för Projektet Nordlab har en lista över vilka kunskaper man måste besitta för att förstå genetik tagits fram (Andersson, 2003c):
1. Alla levande organismer består av celler. 2. Varje cell innehåller kromosomer.
3. Gener är delar av kromosomerna, och innehåller den genetiska informationen.
4. Gener består av DNA. 5. DNA kan kopiera sig själv. 6. Gener kodar för proteiner.
7. Gener kan sättas på och stängas av.
8. Celler kan bli olika beroende på vilka gener som är aktiva. 9. Kromosomer förekommer i par, en från varje förälder. 10. Gener finns i olika varianter.
11. Gener förekommer i par, men det kan vara olika varianter i paret. 12. Vid vanlig celldelning (mitos) bildas celler med samma genetiska
information som modercellen.
13. Alla kroppsceller i organismen innehåller samma genetiska information (alltså identiska kromosomer och gener).
14. Könsceller bildas genom reduktionsdelning (meios) och får därmed hälften av modercellens kromosomer.
15. Könscellerna blir olika, eftersom de får ett urval av förälderns kromosomer. 16. Vid befruktning sammansmälter två könsceller så att antalet kromosomer
återställs.
17. Avkommor ärver genetisk information från sina föräldrar, men blir unika beroende på könscellernas slumpmässiga olikhet.
Denna lista bör ses som ett minimum för vad elever bör känna till efter att ha blivit undervisade i genetik. Dessa punkter är betydligt mer utförliga och konkreta än de mål som finns presenterade i kursplanerna. Det finns inget i den målbeskrivning som togs fram i Projektet Nordlab som motsäger kursplanernas mål och vice versa.
3.5 Hur har läroböcker förändrats över tiden?
Med datorns intrång vid framställning av tryckmaterial har möjligheterna till snygg modern layout ökat. Nya tryckmetoder har ökat möjligheterna och minskat kostnaderna för att använda fyrfärgsbilder vid tryckning.
Skillnaderna mellan nyare och äldre läroböcker begränsas inte till dessa ytliga förändringar. Människors värderingar och vad som anses vara viktigt för elever att kunna, förändras också över tiden. Sådana förändringar kommer att speglas i läroböckerna.
En annan skillnad mellan äldre och nyare böcker som också har noterats i forskningslitteraturen är att den senare gruppen ofta använder ett mer vardagsbetonat språk och enklare förklaringar än den förra gruppen (Ohlsson, 2005). Denna ”snuttifiering” har även noterats i andra ämnen än de naturvetenskapliga (Selander, 1988). En annan trend som har noterats är att den förklarande texten i nyare böcker har kortats ner jämfört med äldre böcker samtidigt som fackuttrycken har behållits (Hägerfelth, 2004).
4. Material och metod
4.1 Val av metod
Några av de vanligaste metoderna för datainsamling i forskningssammanhang är intervjuer, enkäter och dokumentanalys.
Enkäter och intervjuer bygger båda på frågor och har därmed vissa likheter med varandra (Patel & Davidsson, 2003). Vid intervjuer ställer intervjuaren frågor direkt till intervjupersonen. Detta kan ske genom att de båda personerna träffas eller t.ex. talar i telefon med varandra. Med enkäter menas svarsformulär som ska fyllas i. Sådana kommer ofta med posten, men vid vissa enkäter kan personen som utformat enkäten finnas till hands för att kommentera eventuella oklarheter.
Det finns två viktiga parametrar att beakta vid intervjuer, nämligen graden av standardisering och strukturering (Patel & Davidsson, 2003). Med graden av standardisering menas i vilken mån intervjuaren kan påverka frågornas utformning och inbördes ordning. Helt standardiserade intervjuer kan skrivas ner och påminner då till sitt upplägg om enkätundersökningar. Sådana undersökningar används ofta i sammanhang då man vill kunna jämföra olika fenomen och generalisera. Med graden av strukturering menar man i vilken mån intervjupersonen är fri att tolka innebörden av frågorna. Inom en helt strukturerad intervju finns det bara ett fåtal möjliga svarsalternativ.
Enkäter och intervjuer är beroende av personer som är villiga att svara på de frågor som ställts i undersökningarna. Det är därför viktigt att motivera de personer man vill få svar från, annars finns det risk att man får en låg svarsfrekvens (Patel & Davidsson, 2003).
Vid dokumentanalys undersöker man tryckta källor (Patel & Davidsson, 2003). Exempel på dokument är statistik och register, officiella handlingar, privata handlingar, litteratur, tidningar och bilder. Dokument används i undersökningar som vill fastslå faktiska förhållanden eller faktiska skeenden eller individers upplevelser av ett fenomen. Det är vid den här typen av undersökningar viktigt att förhålla sig kritisk till källan. Det kan också i vissa fall vara viktigt att fastställa huruvida källan är original
eller en förfalskning. Hur mycket material man samlar in vid den här typen av undersökningar är beroende av problemställningen samt hur mycket tid man har för att genomföra analysen. Vid analyser av läromedel undersöker man oftast hur språket används, hur olika fenomen förklaras samt vilka fakta författaren har valt att presentera (Selander, 1988)
Syftet med den här studien är att undersöka i vilken mån naturvetenskapliga läroböcker är skrivna på ett sätt som kan tänkas överbrygga de svårigheter som identifierats i den ämnesdidaktiska forskningen. Eftersom jag framförallt är intresserad av hur problemen hanteras i läroböckerna, snarare än hur läroboksförfattarna upplever att de hanteras, valde jag att analysera läroböcker snarare än att använda mig av intervjuer eller enkäter. Jag valde att göra en komparativ, d.v.s. jämförande, undersökning av de olika läroböckerna för att undersöka hur de skiljer sig åt med avseende på hur de hanterar kända svårigheter (Johansson & Svedner, 2001). En fördel med att använda enkäter riktade till läroboksförfattare skulle annars vara att de sannolikt skulle ge snabbare resultat än tidskrävande läroboksanalyser. En nackdel är att informationen man erhåller från enkätstudier tenderar att vara bred, men samtidigt sakna det djup som andra metoder ger (Johansson & Svedner, 2001). Jag bedömde att det var viktigt att tränga in på djupet för att få svar på frågeställningarna i den här studien och valde därför att inte använda mig av enkäter. Kvalitativa intervjuer ger djupare information och skulle kunna vara aktuella för uppföljande framtida studier.
4.2 Avgränsningar och urval
Att bedriva detta projekt inom alla naturvetenskapliga ämnen och läroböcker skulle naturligtvis vara ett monumentalt arbete. Jag har därför valt att avgränsa arbetet till att omfatta genetikavsnittet inom ramen för gymnasieskolans Biologi A. Anledningen till att jag valt denna avgränsning är att genetikavsnittet ofta behandlas väl avskilt från de övriga biologiavsnitten i läroböcker, vilket underlättar jämförelser mellan dessa. Dessutom har genetikämnet identifierats, jämte vattentransport i växter, som det för eleverna svåraste ämnesområdet inom biologi (Bahar et al., 1999). De flesta ämnesdidaktiska studierna är gjorda på den klassiska genetiken och på genetiska processer, varför genteknikavsnitten utelämnats i analyserna.
Jag har bestämt mig för att använda två nya och en äldre lärobok i arbetet:
1. Spira –Biologi A av Gunnar Björndahl, Birgitta Landgren och Mikael Thyberg.
Stockholm: Liber AB 2007.
2. Biologi A med naturkunskap av Janne Karlsson, Thomas Krigsman,
Bengt-Olov Molander och Per-Olof Wickman. Stockholm: Liber AB 2005.
3. Biologi för gymnasieskolan av Yngve Bernhardsson, Sven-Anders Björse och
Arne Müntzing (redaktörer). Esselte Studium 1981.
Vid valet av de båda nyare läroböckerna var den avgörande faktorn publiceringsdatumet. Ju nyare boken är desto mer tid har förflutit sen de ämnesdidaktiska forskningsarbetena publicerades och sannolikheten för att läroboksförfattarna känner till dessa har ökat. De båda nya böckerna var de två senast publicerade biologiböckerna jag kunde hitta vid studiens start. För att läroboksförfattarna ska ha haft en rimlig chans att tillgodogöra sig forskningen bedömer jag att det bör ha förflutit åtminstone tre år mellan forskningspublikationen och lärobokspublikationen.
Den äldre boken valdes så att forskningsresultaten inte ska ha varit kända när boken publicerades. Ifall jag bedömer att de nyare böckerna inte har inkorporerat de senaste vetenskapliga rönen kan jag jämföra med den gamla boken för att se om det har skett någon förändring över tiden.
Jag har alltså valt att använda mig av en komparativ undersökning (Johansson & Svedner, 2001). Den urvalsmetod jag använder mig av är ändamålsenlig på så vis att jag valt böcker tryckta efter ett visst datum (Hartman, 2004). Dessutom är det ett bekvämlighetsurval eftersom jag valde bland de böcker jag har tillgång till. Jag har i upplägget av det här arbetet försökt använda mig av en s.k. logisk disposition (Johansson & Svedner, 2001).
4.3 Om böckerna
Nedan följer en kort beskrivning av de tre läroböcker som användes i den här studien, samt information om vilka delar som användes i analysen.
4.3.1 Spira –Biologi A
Spira är den senast publicerade läroboken i den här studien. Boken omfattar 266 sidor
och är anpassad för gymnasieskolans Biologi A-kurs (Björndahl et al., 2007). Den har en modern layout och är rikligt illustrerad med både förklarande bilder och intresseväckande fotografiska tryck. Spira är uppdelad i fyra block med vardera några kapitel. Blocken är: I: Biologi handlar om liv, II Genetik, III Livets mångfald och IV
Samspel i naturen. Det första blocket behandlar bl.a. grundläggande information om
biologi, vetenskapsteori, celler och biologiska organisationsnivåer. Det andra blocket behandlar genetik och genteknik medan det tredje behandlar systematik och evolution. Det sista blocket handlar om ekologi.
Kapitlen varierar i längd mellan ett par sidor upp till strax under 20 sidor och avslutas med en sammanfattning. I slutet av boken finns ett antal övningsuppgifter på vart och ett av de olika kapitlen.
Genetikavsnittet ryms i block II, och kommer därmed relativt tidigt i boken. Block II omfattar de fem kapitlen: 3. DNA och RNA; från molekyl till individ (s. 34-42), 4.
Kromosomer, celldelning och förökning (s. 43-50), 5. Mutationer (s. 51-58), 6. Klassisk genetik (s. 59-75) och 7. Genteknik (s. 76-93). I den här studien har jag valt att analysera
kapitel 3-6, vilka omfattar totalt 42 sidor. Av dessa 42 sidor utelämnades 4 då de var allmänna inledningar eller sammanfattningar.
Spira har en egen webbplats med ytterligare övningar och sammanfattningar på
adressen: www.liber.se/spira.
4.3.2 Biologi A med naturkunskap
Biologi A med naturkunskap är anpassad för gymnasieskolans Biologi A- och
Naturkunskap A-kurser och är 310 sidor tjock (Karlsson et al., 2005). Den har liksom
indelad i nio delar, vart och ett med ett antal kapitel. De olika delarna är: Ekologi,
Hållbar utveckling, Cellens byggnad, Cellens genetik, Individens genetik, Genteknik, Evolution, Etologi och beteendeekologi samt De levande organismerna. Flera av
delarna innehåller vissa avsnitt som bör läsas inom ramen för biologi och andra avsnitt som är avsedda som en del av naturkunskapsundervisningen. De olika avsnitten är tydligt utmärkta.
Varje kapitel innehåller från ett par sidor upp till strax över tio sidor och avslutas med ett antal frågor och en sammanfattning i form av en mindmap.
Genetikämnet behandlas i de tre delarna Cellens genetik (s. 143-166), Individens genetik (s. 167-188) och Genteknik (s. 189-206). Liksom i Spira avhandlas alltså genetiken relativt tidigt i boken. I den här studien har jag valt att analysera de första båda kapitlen. Jag har dessutom valt att välja bort kapitlen Virus är proteinklädda nukleinsyror (s. 160-161) och Hur vet man att DNA bär på arvet? (s.162-164) i Cellens genetik, då jag bedömer att dessa i det förra fallet inte behandlar genetikämnet och i det senare fallet snarare handlar om en genteknisk metod. Totalt omfattar materialet 41 sidor, varav 6 sidor utelämnades då dessa bestod av allmänna inledningar eller sammanfattningar.
Biologi A med naturkunskap har en egen webbplats med ytterligare övningar och
sammanfattningar på adressen: www.liber.se/gymnasiebiologi.
4.3.3 Biologi för gymnasieskolan
Biologi för gymnasieskolan är den tidigast publicerade boken i den här studien. Boken
som har underrubriken Genetik, Tillväxt och differentiering, Evolution och Etologi är en del i en serie om tre böcker som var avsedda för gymnasieskolans naturvetenskapliga linje och den frivilliga undervisningen i biologi på teknisk linje (Bernhardsson et al., 1981). Boken omfattar 188 sidor och har en betydligt äldre layout än de andra båda böckerna i den här studien. Liksom de båda nyare böckerna har den rikligt med bilder om än svartvita. Illustrationerna är plottrigare än i de båda nyare böckerna, sannolikt för att de är ritade för hand och inte med hjälp av dator som i de nyare böckerna. Sammantaget ger Biologi för gymnasieskolan ett betydligt tråkigare intryck än andra båda böckerna.
Som bokens underrubrik antyder innehåller boken fyra delar: Genetik, Tillväxt och
differentiering, Evolution och Etologi. Var och en av delarna innehåller ett antal kapitel
som omfattar mellan ett par upp till strax under 15 sidor. Varje kapitel avslutas med en kort sammanfattning. Boken innehåller inga övningsuppgifter och har av lättförklarliga skäl ingen webbplats.
Genetikämnet avhandlas i inledningen av boken, och finns representerat i de båda delarna Genetik (s. 1-72) och Tillväxt och differentiering (s. 73-105). Samtliga 13 kapitel under den förra delen, samt de tre första kapitlen i den senare delen har analyserats i den här studien. Med utelämnade allmänna inledningar och sammanfattningar omfattade det analyserade materialet 78 sidor.
4.4 Metod
Vid analyserna av läroböckerna utgick jag från Knippels (2002) lista över vanliga problem inom genetikundervisningen, vilka presenterades i 3.3 Didaktiska problem
inom genetikundervisning:
1. Ämnet innehåller många fackuttryck.
2. Matematiska beräkningar inom ämnet kan vara komplicerade. 3. Cellulära processer kan vara svåra att förstå.
4. Ämnet kan bli abstrakt beroende på undervisningens upplägg. 5. Snabba hopp mellan makro- och mikronivå kan vara förvirrande.
4.4.1 Ämnet innehåller många fackuttryck
Läroböckerna analyserades med avseende på antalet fackuttryck, antalet fackuttryck per sida samt antalet nya fackuttryck per sida. Resultaten från denna analys skulle kunna indikera om onödigt många facktermer används. Om en fackterm endast används vid ett eller ett par tillfällen kan man ifrågasätta meningen med att introducera termen. Jag valde att inte bara inkludera rent genetiska termer bland de räknade fackuttrycken, utan även andra biologiska termer, namn på kemikalier och latinska artnamn. Facktermerna behandlades alltså kvantitativt, i en analys som var närmast induktivt positivistisk till upplägget (Hartman, 2004).
4.4.2 Matematiska beräkningar inom ämnet kan vara komplicerade
Analyserna av de övriga problemen var mer subjektivt utformade och innehåller försök att tolka läroboksförfattarnas intentioner och hur väl dessa fallit ut. Dessa analyser bygger på en hermeneutisk vetenskaplig grund (Hartman, 2004). Det kan kanske tyckas motsägelsefullt att använda både positivistiska och hermeneutiska metoder i samma studie, men detta är inget ovanligt fenomen. I själva verket vet många forskare inte vilken av de båda skolbildningarna de arbetar efter (Patel & Davidsson, 2003).
För att undersöka hur läroböckerna behandlar matematiska beräkningar inom genetikämnet, undersöktes om böckerna hade någon inledande repetition av sannolikhetsberäkningar. Att inleda med beräkningar av vardagsproblem skulle kunna underlätta för eleverna att få in rätt sätt att tänka vid genetiska sannolikhetsberäkningar. Dessutom undersöktes texterna för att se om symboler användes på ett konsekvent och logiskt vis vid t.ex. korsningsscheman och släktträd. Vid förekomst av korsningsscheman undersöktes också om texterna underlättade för eleverna att inte tolka dessa deterministiskt.
4.4.3 Cellulära processer kan vara svåra att förstå
Vid analyserna av detta problem undersöktes hur väl meiosbegreppet förklaras, och huruvida skillnader mellan meios och mitos tydliggörs i texterna. Vidare undersöktes om mitos och meios avhandlas direkt efter varandra i texterna samt huruvida kromatidbegreppet används eller inte.
4.4.4 Ämnet kan bli abstrakt beroende på undervisningens upplägg
Texterna analyserades med avseende på om meiosen behandlas i samband med genetiken samt huruvida detta begrepp kopplas ihop med den klassiska genetiken. Dessutom undersöktes om mitos och meios kopplas ihop med cellcykeln samt huruvida separata cellinjer presenteras för de båda fenomenen.
4.4.5 Snabba hopp mellan makro- och mikronivå kan vara
förvirrande
Avslutningsvis analyserades hur hopp mellan olika storleksdimensioner hanteras i de tre läroböckerna. Undviks onödiga hopp? Förklaras fenomen på båda makro- och
mikronivå? Inleds förklaringarna på makronivå och fortsätter successivt till lägre nivåer?
4.5 Tillförlitlighet och generaliserbarhet
Syftet med den här studien var att ta reda på om den senaste didaktiska forskningen tas in i nya läroböcker. För att undersöka detta använde jag mig av två aktuella biologiböcker i vilka genetikavsnitten analyseras. En begränsning med det här upplägget är naturligtvis att textanalyserna bara omfattar en bråkdel av de läroböcker som finns i gymnasieskolan. Det är inte utifrån den här studien möjligt att entydigt svara på frågan om läroboksförfattaren generellt sett lyssnar på forskaren eller inte. Däremot bör studien kunna svara på om de läroboksförfattare som har skrivit de analyserade kapitlen har använt sig av den ämnesdidaktiska forskningen eller inte.
Ett potentiellt problem med den här studien är att det mesta av den ämnesdidaktiska forskningen har utförts i andra länder än Sverige, vilket öppnar för möjligheten att de svårigheter som de utländska barnen har i genetik kanske inte är desamma som dem svenska barn har. Dessutom är studierna ibland gjorda på elever som är något yngre än svenska gymnasieelever i regel är när de läser genetik.
Ytterligare ett potentiellt problem i min studie är att de problem som presenteras i Knippels (2002) inte är specifikt anpassade för läroböcker utan gäller allmänt vid undervisning i genetik. Kanske är en del av problemen svåra att direkt applicera på en lärobok?
Vid analysen av facktermer kan jag ha missat några ord. Dessutom är det inte självklart vad som fick definiera en fackterm. Jag inkluderade förutom rena genetiktermer en del knepiga sammansatta ord, latinska artnamn samt andra biologiska termer. Till exempel valde jag att ta med äggcell men inte spermie. Anledningen till detta är att flera andra sorters celler tas upp och jag anser att dessa lätt kan blandas ihop.
5. Resultat
Nedan följer resultaten för läroboksanalyserna. Resultaten för alla tre böckerna presenteras tillsammans för vart och ett av de fem problemområden som analyserna utgick ifrån (se 4. Material och metod).
5.1 Ämnet innehåller många fackuttryck
Antalet facktermer beräknades enligt 4. Material och metod. Resultaten redovisas i sin helhet i Bilaga 1 och som en sammanfattning i Tabell 5.1. Dessutom presenteras de samlade resultaten från läroboksanalysen i Tabell 5.2. Som framgår av Tabell 5.1 använder alla de analyserade böckerna många fackuttryck. Totalt används strax under 200 facktermer i de båda nyare böckerna, medan över 200 facktermer används i den äldre boken. Om man tar hänsyn till antalet sidor i analyserna ser man att Björndahl et
al. (2007) och Bernhardsson et al. (1981) använder betydligt färre fackuttryck per sida
än Karlsson et al. (2005). Om man däremot beräknar antalet nyintroducerade termer per sida finner man att de båda nyare läroböckerna kommer upp i 5 termer på sida medan den äldre boken bara innehåller 3 nya termer per sida. Trots att Karlsson et al. (2005) använder betydligt fler facktermer per sida än Björndahl et al. (2007), introduceras alltså inte fler nya ord per sida. I den senare boken används de introducerade orden i lägre frekvens än i den förra.
Tabell 5.1. Sammanfattning av resultaten från fackuttrycksberäkningen.
Björndahl et al. 2007 Karlsson et al. 2005 Bernhardsson et al. 1981 Analyserade sidor 38 35 79 Antal fackuttryck 186 172 219
Fackuttryck per sida 21 37 16
Björndahl et al. (2007) innehåller 106 facktermer som används färre än tre gånger medan motsvarande siffror för Karlsson et al. (2005) och Bernhardsson et al. (1981) är 73 respektive 115.
Alla tre böckerna använder sig av synonymer bland facktermerna, vilket gör att antalet ökar. Till exempel använder alla böckerna termerna budbärar-RNA och messenger-RNA parallellt (Bilaga 1). Andra exempel är att arvsmassa och arvsubstans används synonymt med DNA i Björndahl et al. (2007) respektive Bernhardsson et al. (1981). I Karlsson et al. (2005) används termen systerkromatid parallellt med termen identiska
kromatider. Alla läroböckerna använder sig av begreppet kromatid (i Björndahl et al.
(2007) används endast termen systerkromatid).
Böckerna innehåller många ord som liknar varandra och därmed skulle kunna blandas ihop. I Karlsson et al. (2005) används t.ex.: kromosom-kromatid, gen-genom-genotyp,
homolog-homozygot, ribos-ribosom, transkription-translation-transfer RNA, centromer-centrosom.
Synonymer: danssjuka-Huntingtons sjukdom, meios-reduktionsdelning,
styrsekvens-promotor, startsekvens-primer, mitos-vanlig celldelning, identiska kromatider-systerkromatider.
I Björndahl et al. (2007) används: kromosom-kromatid, gen-genom-genotyp,
homolog-homozygot, ribos-ribosom, transkription-translation-transfer RNA, centromer-centrosom m.fl.
Motsvarande lista i Bernhardsson et al. (1981) lyder:
allel-alloploidi-allopolyploidi-allotetraploid, anafas-aneuploid, autoploidi-autopolyploidi-autotetraploid, cellulosa-centriol-centromer, cytoplasma-cytosin, diplofas-diploid, dottercell-dotterkromosom-dottercentriol, fenylalanin-fenylketonuri-fenylpyrodruvsyra, gen-genom-genotyp m.fl.
Böckerna innehåller också ett antal felaktiga uppgifter och formuleringar som skulle kunna försvåra för läsaren att tillgodogöra sig materialet. Nedan följer några exempel på sådana felskrivningar och dunkla formuleringar.
Ur Spira (Björndahl et al., 2007):
• ”DNA-molekylen är både större och mer komplex än RNA” (s. 36). Detta är en tveksam formulering. RNA- och DNA-molekyler har ytterst små strukturella skillnader och är således ungefär lika komplexa. Storlekarna på molekylerna varierar, men båda ämnena förekommer i både stora och små former.
• ”DNA består av två nukleotidkedjor med en ryggrad av fosfat och sockret
deoxiribos” (s. 36) Detta är också en formulering som inte är helt korrekt. DNA
består av en nukleotidkedja. Dubbelsträngat DNA består av två kedjor, men fosfat- och sockergrupperna är en del av nukleotiderna. Så som meningen nu är formulerad låter det som om DNA förutom nukleotider också består av socker och fosfat, vilket alltså inte är fallet.
• Kvävebaser förkortas på flera ställen till baser och på andra ställen används de felaktigt som synonyma med nukleotider.
• Vid en förklaring av ett korsningsschema står: ”När man bara tittar på
plantorna går det inte att avgöra vilka som är homozygota respektive heterozygota” (s. 63). Men på bilden över blommorna återges de i olika färger.
Förmodligen avses att man inte ser skillnad på de dominant homozygota och heterozygota fenotyperna.
Ur Biologi A med naturkunskap (Karlsson et al., 2005):
• ”[…] dominanta och recessiva egenskaper […]” (s. 174) används istället för dominanta och recessiva alleler. Detta misstag återkommer på flera ställen i texten.
• Termen ”startsekvens” används ibland för att indikera startpunkten för translationsprocessen och ibland synonymt med termen ”primer”, vilket är något helt annat.
• Termen ”kopplade egenskaper” används istället för kopplade gener. Ibland talas det om ”kopplade kromosomer” istället för kopplade gener (s. 176).
• Spermier som bär på X-kromosomer kallas ”X-spermier” (s. 179).
• Texten talar om överkorsning mellan två ”loci” istället för mellan två
kromosomer (s. 176).
• Termen ”autosomal allel” används istället för ”allel på en autosomal
kromosom” (s. 182).
Ur Biologi för gymnasieskolan (Bernhardsson et al. 1981):
• Vid beskrivning av en befruktning talas om ”hankärnor” och ”honkärnor” istället för cellkärna från hane respektive hona (s. 1).
• "Man vet numera med säkerhet att arvsubstansen hos alla organismer består av
nukleinsyror" (s. 4). Att inom naturvetenskapen säga att man vet någonting med
säkerhet är felaktigt. Naturvetenskaplig vetenskapsteori bygger på teorier formulerade utifrån hypoteser.
• Termen ”nukleotidbas” används istället för ”kvävebas” (s. 4).
• Termerna ”nykombination”, ”omkombination” och ”rekombination” används i texten med olika betydelser. Att använda så lika ord för olika processer kan bli förvirrande för läsaren.
• Begreppet ”gen” används på flera ställen där allel avses. T.ex. ”genpar” (s. 18),
• I allelnamn representeras ibland dominanta och recessiva alleler med andra beteckningar än stora respektive små bokstäver (t.ex. på s. 51).
• Människor med en X-kromosom och två Y-kromosomer kallas
”XYY-människor” (s. 54). Detta kan få det att låta som om hela människan definieras
av en mutation i cellerna.
De tre läroböckerna använder alla fackuttryck som även används i andra sammanhang. Exempel på detta är termen dominant, som används om alleler. I Karlsson et al. (2005) och Bernhardsson et al. (1981) markeras inte att denna term har en annan innebörd inom genetikämnet än i vardagssammanhang. I Björndahl et al. (2007) markeras detta tydligt: ”Själva begreppet dominant betyder bara att allelen visar sig i enkel upplaga,
inte på något vis att den är vanligare eller bättre” (s. 63).
5.2 Matematiska beräkningar inom ämnet kan vara
komplicerade
De tre analyserade läroböckerna använder alla korsningsscheman och sannolikhetsberäkningar. Inga av böckerna innehåller någon repetition av allmänna sannolikhetsberäkningar innan de genetiska sannolikheterna presenteras. Författarna markerar inte heller på ett tydligt sätt att korsningsscheman inte är deterministiska eller diskuterar deras begränsningar. I Karlsson et al. (2005) står om korsningsscheman att:
”I ett sådant schema kan föräldrarnas alleler kombineras, så att man får reda på avkommans allelkombinationer” (s.172). I ett korsningsschema får man reda på
avkommans möjliga allelkombinationer och sannolikheten för att var och en av dem ska uppträda i avkomman.
Vid beskrivningar av korsningsscheman bör dessa kopplas ihop med meiosen och dess slumpvisa segregering av gameter (Cho et al., 1985). Detta sker i Karlsson et al. (2005) (s. 171) och Bernhardsson et al. (1981) (s. 13) men inte i Björndahl et al. (2007).
I Karlsson et al. (2005) används allelbeteckningar på ett konsekvent sätt: dominanta alleler markeras med stora bokstäver medan recessiva alleler markeras med små
bokstäver. I de övriga båda böckerna gör författarna avsteg från detta system. I Bernhardsson et al. (1981) används ibland beteckningarna I1-I5 (s. 61), medan Björndahl
et al. (2007) använder beteckningarna XHY, XHXh m.fl. (s. 69).
5.3 Cellulära processer kan vara svåra att förstå
I Bernhardsson et al. (1981) beskrivs inte mitos och meios direkt efter varandra som i de båda nyare böckerna. Däremot kopplas inte dessa båda celldelningar ihop med övriga cellcykeln, vilket är fallet för de båda nyare böckerna.
Alla de analyserade böckerna behandlar mitos och meios inom ramen för genetikämnet, men inga av dem markerar tydligt skillnaderna dem emellan.
5.4 Ämnet kan bli abstrakt beroende på undervisningens
upplägg
Celldelningen behandlas före den klassiska genetiken i alla de analyserade läroböckerna. Dessa till varandra kopplade fenomen binds inte heller ihop i texterna utan förblir separata från varandra. I de båda nyare böckerna beskrivs cellcykeln före celldelningsavsnittet. I dessa böcker introduceras också kromosombegreppet före celldelningen medan endast Björndahl et al. (2007) introducerar kromatidbegreppet före celldelningen. I den senare boken kommer dock kromatidbegreppet även före beskrivningen av cellcykeln. Bernhardsson et al. (1981) beskriver mitosen separat från celldelningen och före introduktionen av de båda termerna kromosom och kromatid. Böckerna beskriver könsceller och somatiska celler som separata cellinjer, men bara vid själva celldelningen, inte vid beskrivningen av cellcykeln.
Som beskrevs ovan använder bara Karlsson et al. (2005) allelbeteckningar på ett konsekvent sätt. Vad gäller beteckningar i släktträd använder de olika böckerna ett konsekvent symbolspråk inom respektive bok. Bara Karlsson et al. (2005) använder den gängse metoden att markera endast fenotypiska skillnader i stamträden. Kvinnor markeras med cirklar medan män representeras av kvadrater. Sjuka individer markeras med fylld symbol. Björndahl et al. (2007) och Bernhardsson et al. (1981) markerar
dessutom anlagsbärare. I den senare boken markeras dessa med en färgad prick i mitten av cirkeln eller kvadraten och i den tidigare boken markeras de med en halvfylld symbol.
5.5 Snabba hopp mellan makro- och mikronivå kan vara
förvirrande
Samtliga av de analyserade böckerna inleder genetikavsnitten på cellulär eller molekylär nivå. De båda nyare böckerna inleder med nukleinsyrans uppbyggnad och går sedan via genbegreppet vidare till transkription, translation och proteinsyntes. Karlsson et al. (2005) behandlar sedan den klassiska genetiken medan Björndahl et al. (2007) först beskriver olika sorters mutationer. Bernhardsson et al. (1981) inleder med mitosen och går därefter in på nukleinsyrans struktur. Därefter beskrivs meiosen följd av den klassiska genetiken.
Läroböckerna hoppar inte i nämnvärd grad mellan makro- och mikronivåer. Vid beskrivningar av blomfärger och liknande, nämns allelbeteckningarna i samband med blommans färg, men det torde vara nödvändigt för förståelsen att använda hopp i detta sammanhang.
5.6 Sammanfattning av resultaten
Resultaten i den här studien sammanfattas i Tabell 5.2. nedan. Analysresultaten från de tre böckerna redovisas med avseende på 19 parametrar. Dessutom redovisas rekommendationer från forskningslitteraturen för hur de olika delarna bör behandlas.
Tabell 5.2. Sammanfattning av analysresultaten från de tre läroböckerna.
Innehåller texten… Björndahl Karlsson Bernhardsson Enl. litt. Referens
många fackuttryck? ja (186) ja (172) ja (219) undvikes Bahar et al. 1999 många komplexa termer? ja ja ja undvikes Bahar et al.
många liknande termer? ja ja ja undvikes Bahar et al. 1999 många synonymer? ja ja ja undvikes Bahar et al.
1999 felanvända termer? ja ja ja nej Cho et al.
1985 extra förklaring av termer
som även används i vardagsspråk?
ja nej nej ja Pearson & Hughes
1988 klargörande att
korsningsscheman ej är deterministiska?
nej nej nej ja Collins & Stewart 1989 beskrivning av slumpvis segregering av gameter vid korsningsscheman? nej ja ja ja Kinnear 1983 repetition av sannolikhetslära i samband med beräkningar?
nej nej nej ja Kinnear 1983
mitos och meios i
samband med cellcykeln?
ja ja nej ja Cho et al. 1985
tydliga markeringar av skillnader mellan mitos och meios?
nej nej nej ja Cho et al. 1985
kromatidbegreppet? ja ja ja undvikes Longden 1982
meiosen i samband med genetikavsnittet?
ja ja ja ja Cho et al. 1985
koppling mellan meiosen och den klassiska genetiken?
nej nej nej ja Cho et al. 1985
meiosen och mitosen direkt efter varandra?
ja ja nej undvikes Bahar et al. 1999
konsekvent användning av symboler?
nej ja nej ja Bahar et al. 1999 koppling mellan korsningsscheman och biologiska fenomen? ja ja ja ja Thompson & Stewart 1985
genetiska fenomen först på makronivå och sedan på mikronivå?
nej nej nej ja Marbach-Ad & Stavy
2000 hopp mellan mikro- och
makronivå i nämnvärd grad?
nej nej nej undvikes Bahar et al. 1999
6. Diskussion
Skillnaderna mellan de analyserade läroböckerna är slående. De nyare båda böckerna har en modernare layout, med tydligare och snyggare bilder. De har intresseväckande färgfotografier och ett modernt språk. Skillnaderna begränsar sig dock inte till dessa ytliga kriterier utan går djupare än så. Det material som författarna valt att ta med skiljer sig mellan de nyare böckerna å ena sidan och den äldre boken å den andra. I den senare boken ligger fokus mer på växtförädling och djuravel än i de förra. De molekylära processerna beskrivs noggrannare i de nyare böckerna och genteknikavsnitten i dessa böcker är större än i Bernhardsson et al. (1981). Denna bok innehåller flera passager som numera förmodligen skulle klassas som politiskt inkorrekta och utelämnas ur en lärobok. Sålunda beskrivs de olika människoraserna på följande sätt i Bernhardsson et
al. (1981):
”Utom den europida rasen består världens befolkning i huvudsak av mongolida och negroida raser.” (s. 46)
I texten beskrivs sedan olika undergrupper till dessa raser och på bilder återges porträtt av människor från olika raser på ett stereotypt sätt. I denna bok visas också bilder på nakna människor med olika former av genetiska sjukdomar, något som saknas i de nyare böckerna. Som kontrast visas i Karlsson et al. (2005) en bild på en flicka med Downs syndrom som leende leker med andra barn (s. 181). I bildtexten betonas vikten av stimulans för den personliga utvecklingen hos flickan.
Det är intressant att notera att de nyare böckerna bara använder hälften så många sidor för att beskriva samma delar inom genetiken som Bernhardsson et al. (1981) (Tabell
5.1). Den här sortens ”snuttifiering” av läroböcker har noterats vid läromedelsanalyser i
andra ämnen (Selander, 1988). Kanske tänker sig läromedelsförfattarna att det blir lättare att ta till sig en text om den inte är så omfattande. Samtidigt skulle en alltför kortfattad text kunna leda till att det blir svårt för eleverna att se någon helhet i ämnet. Det är intressant att notera att Bernhardsson et al. (1981) använder sig av färre fackuttryck per sida än de båda nyare böckerna. Detta gäller både för det totala antalet fackuttryck per sida och för antalet nyintroducerade fackuttryck per sida (Tabell 5.1).
Det verkar alltså som om författarna till de nyare böckerna framförallt skär ner på den förklarande texten medan antalet fackuttryck behålls. Denna tendens att behålla fakta men att skära ner den berättande texten har även noterats i andra undersökningar (Hägerfelth, 2004). Ur ett socialkonstruktivistiskt perspektiv skulle detta kunna leda till att eleverna får svårare att konstruera sin kunskap eftersom diskussionen enligt detta synsätt är en viktig komponent i lärandet (Andersson, 2001). Alla tre böckerna använder ett stort antal fackuttryck. Så här lyder t.ex. en mening i Bernhardsson et al. (1981):
”En eljest normal kornplanta har genom mutation blivit heterozygot för två recessiva gener. I avkomman från en sådan normalgrön AaBb-planta får man dihybrid klyvning i 9 grön (homo- eller heterozygot för generna A och B), 3 ljusgrön (recessiv homozygot aa), 3 med spår av klorofyll (recessiv homozygot bb), 1 helvit (recessiv homozygot)” (s.
31)
Även i de nyare böckerna förekommer liknande, mycket komplicerade redogörelser:
”Kromosomerna flyttas till ett plan mitt i cellen genom att proteintrådarna drar och knuffar dem. I detta ekvatorialplan placeras kromosomerna så att systerkromatidernas centromerer är riktade mot var sin sida av cellen. Sedan dras systerkromatiderna med hjälp av kärnspolen till var sin cellhalva.” (Karlsson et al., 2005, s. 157)
”Under anafasen skiljs systerkromatiderna från varandra och kallas sedan för kromosomer (där varje kromosom nu utgörs av en DNA-molekyl). Parallellt med telofasen skiljs de två dottercellerna från varandra i en process som kallas cytokines”
(Björndahl et al., 2007, s.46)
6.1 Ämnet innehåller många fackuttryck
Det är knappast möjligt att bedriva undervisning i genetik utan fackuttryck. Vissa termer kan dock undvikas. Elever har t.ex. ofta problem med att förstå skillnaden mellan termerna kromatid och kromosom. Eftersom kromatidbegreppet gör väldigt lite för att öka elevernas förståelse för meiosfenomenet bör det utelämnas (Longden, 1982). Likväl används begreppet i alla de undersökta läroböckerna.
Trots att Björndahl et al. (2007) och Karlsson et al. (2005) är upplagda på liknande sätt innehåller den förra 88 facktermer som inte finns med i den senare (Bilaga 1). Alla tre böckerna innehåller många ord som bara förekommer ett par gånger och inte finns med i de andra båda läroböckerna. En hel del av dessa termer skulle med största sannolikhet ha kunnat utelämnas utan att elevernas förståelse hade blivit lidande (kanske snarare tvärtom). Några exempel på sådana ord är: pleiotropi, epistasi, polymera gener samt namn på olika gener som p53, SRY, FOXP2 m.fl. (Bilaga 1).
Björndahl et al., (2007) och Karlsson et al., (2005) är som sagt upplagda på liknande sätt. Genetikavsnitten är ungefär lika långa och innehåller ungefär lika många unika fackuttryck (Tabell 5.1). Däremot skiljer sig böckerna åt när det gäller hur ofta varje fackterm återkommer. Karlsson et al. (2005) använder 37 fackuttryck per sida medan Björndahl et al. (2007) ”bara” använder 21. Att använda en stor andel facktermer i texten kan troligtvis försvåra för eleverna att tillgodogöra sig materialet. En alternativ tolkning skulle kunna vara att eleverna snarare har nytta av att termerna återkommer fler gånger genom att de då vänjer sig vid dem och får dem repeterade för sig. Enligt socialkonstruktivistisk teori skulle kanske ett utökat diskuterande och användande av facktermerna hjälpa eleverna att ”konstruera” kunskap kring fenomenet.
Alla böckerna innehåller termer som liknar andra termer i böckerna. En del av dessa är nog oundvikliga medan andra skulle kunna tas bort. Vi har redan tagit upp begreppen
kromosom och kromatid, där det senare skulle kunna utelämnas. En vanlig term inom
genetiken är även kromatin, vilken inte finns med i någon av böckerna och ytterligare skulle ha kunnat öka komplexiteten. Inte heller begreppen nukleosid, nukleus eller
nukleol finns med i böckerna. Dessa skulle annars lätt ha kunnat blandas ihop med
termen nukleotid.
Ibland tolkar elever dominanta alleler som varande bättre för individen än recessiva
alleler (Pearson & Hughes, 1988). Av de analyserade läroböckerna är det bara i
Björndahl et al. (2007) som detta missförstånd tydliggörs. Den här sortens missförstånd förekommer ofta bland elever som studerar naturvetenskapliga ämnen (Sanders, 1993).
De analyserade böckerna innehåller alla felaktig eller slarvig användning av facktermer. Detta problem har även uppmärksammats inom de andra naturvetenskapliga ämnena (Sanders, 1993). Det är naturligtvis svårt att helt utplåna sådana felaktigheter, men genom att få texten granskad av utomstående experter före publicering skulle nog en hel del av dessa felaktigheter kunna rättas till. Cho et al. (1985) tar upp den felaktiga användningen av termerna gen och allel som varande synonyma. Detta misstag är vanligt även bland experter. Bernhardsson et al. (1981) begår detta misstag trots att en av medförfattarna, Arne Müntzing var professor i genetik i Lund (Müntzing, 1977). Att använda facktermer på detta felaktiga sätt försvårar för elever att ta till sig materialet (Cho et al., 1985). Inom socialkonstruktivistisk teori är samtalet och diskussionen en viktig komponent i lärandet (Andersson, 2001). När begrepp används på ett felaktigt sätt skulle detta kunna försvåra för eleverna att ”konstruera” kunskap. Karlsson et al. (2005) talar om recessiva egenskaper istället för recessiva alleler. Begreppen allel och
gen används korrekt men i kapitlet ”cellens genetik” används bara termen gen medan
termen allel nästan uteslutande används i kapitlet ”individens genetik”. Detta riskerar att göra det svårare för läsaren att förstå skillnaden mellan begreppen. Att elever missförstår kursstoffet har identifierats som ett vanligt didaktiskt problem inom undervisning i de naturvetenskapliga ämnena (Sanders, 1993).
Det torde nu stå klart att de tre böckerna bara i liten mån tagit till sig av de rekommendationer som den ämnesdidaktiska forskningen kommit fram till avseende användandet av facktermer inom genetikämnet. Ingen av böckerna sticker ut genom att vara avsevärt bättre eller sämre beträffande användandet av facktermer. De har alla styrkor och svagheter.
Hindrar gymnasieskolans kursplan författarna från att använda färre facktermer? Målen för gymnasieskolans Biologi A-kurs är mycket generellt angivna, och kolliderar inte på något sätt med de rekommendationer som ämnesdidaktiska forskningen inom genetik har kommit fram till. Målen i kursplanerna både för gymnasieskolan och grundskolan som relaterar till genetik är så kortfattade och generella att det är nödvändigt för lärarkåren att göra egna lokala uttolkningar för att definiera vad de egentligen vill att eleverna ska kunna efter att ha genomgått kursen. Detta får sannolikt till följd att elever på olika skolor går igenom olika delar av genetiken med skiftande djup och noggrannhet.
De ämnesdidaktiska rekommendationerna kolliderar inte heller med den lista med punkter över vad man bör kunna för att behärska genetik som Projektet Nordlab har sammanställt (se 3.4).
6.2 Matematiska beräkningar inom ämnet kan vara
komplicerade
Matematik och sannolikhetsberäkningar har identifierats som problemområden inom genetikundervisningen (Kinnear, 1983). Eleverna behärskar ofta sannolikhetsberäkningar på vardagsfenomen men blir osäkra när problemen är av genetisk karaktär. Ett sätt att råda bot på detta skulle kunna vara att börja med sannolikheter på vardagsnivå och sedan föra in resonemanget på ett genetiskt problem. Inga av böckerna gör på detta vis, utan börjar direkt med sannolikheter på genetiska problem. Detta försvårar sannolikt för eleverna att ta till sig innehållet.
En av orsakerna till att elever upplever sannolikheterna som svåra inom genetikämnet är en förvirrande användning av symboler. Karlsson et al. (2005) är den enda av böckerna som använder allelbeteckningar på ett konsekvent sätt (Tabell 5.2). Denna bok inledningen stycket om klassisk genetik med en kort repetition av meiosen och beskriver den slumpvisa segregationen av homologa kromosomer, vilket sannolikt gör det lättare för läsaren att förstå bakgrunden till korsningsscheman. I den första bilden på ett korsningsschema har dock författarna gjort misstaget att skriva ”hane” och ”hona” på axlarna istället för att tydligt markera att det rör sig om de olika möjliga allelutfallen från meiosen.
Ett annat vanligt problem vid missförstånd av korsningsscheman är att de tolkas deterministiskt (Collins & Stewart, 1989). I Karlsson et al. (2005) markeras inte tydligt att korsningsscheman inte ska tolkas deterministiskt. Däremot presenteras Mendels experiment med ärter på ett sätt så att det framgår att klyvningstalen inte blev exakt som förväntat. Detta borde till viss del hjälpa eleverna att inte tolka korsningsscheman som deterministiska.