Early Holocene Seal
Hunting on the
Island of Gotland
University of Gotland
2011 / Spring term
Bachelor's essay
Author: Alexander Sjöstrand
Department of Culture, Energy and Environment
Osteology
Supervisors: Senior lecturer Carola Liebe-Harkort,
Senior lecturer Jan Apel
Assistant master Margareta Kristiansson
Gisslause during the excavation of summer 2010. Photo by Anders GustavssonAbstrakt
Kandidatuppsats av Alexander Sjöstrand. Högskolan på Gotland.
Early Holocene Seal Hunting on the Island of Gotland - Mesolitisk säljakt på Gotland. Denna studie inkluderar resultat från osteologiska analyser från tre stenålderslokaler på Gotland: Visborgs Kungsladugård, Strå samt Gisslause. Dessa tre lokaler är daterade till Mesolitikum, dvs. bland de tidigaste faserna på Gotland. En majoritet av de identifierade benen har varit från säl och därför har uppsatsen kommit att handla om den säljakt som utövats på dessa tre platser. Utifrån de fynd som analyserats har skillnader kunnat uttolkas vad det gäller både åldersfördelning och fördelning av olika sälarter mellan vissa av lokalerna. Visborgs Kungsladugård visar på en majoritet av yngre gråsälar till skillnad från Strå och Gisslause där det existerar en jämn
fördelning mellan gråsäl och vikare, likaså en jämn fördelning mellan åldrarna. Tolkningen av detta pekar på en delvis säsongsjakt där populationerna har jagat på olika sätt på olika delar av Gotland. I uppsatsen har även en diskussion angående den marina anpassningen tagits upp. Denna anpassning har setts i tre stadier,
lakustrin, littorin och marin. Den mesolitiska populationen på Gotland passar in på det littorina stadiet där en full marin kultur inte ännu varit helt utvecklad. Då det främst jagats säl på och nära stranden samt att ett visst insjöfiske utövats påvisar resultaten att ett fullt utnyttjande av havet ännu inte bedrivits.
Acknowledgements
I would like to thank Carola Liebe-Harkort for her dedication when helping with the essay, both with the osteological material as well as with the writing and editing of the text. I would also like to extend a large thanks to Jan Apel for introducing me into this project as well as letting me be a part of it, both in analysis and excavation and supplying me with ideas and literature. Additionally I would like to thank Jan Storå for helping me with seal method and other questions regarding seal. Last but not least I would like to thank Margareta Kristiansson for helping with the osteological analysis.
Index
1.1 Island fauna... 2
1.2 Earlier research ... 4
2. Aim and Purpose ... 4
3. Material ... 5
3.1 Visborgs Kungsladugård ... 5
3.2 Strå ... 5
3.3 Gisslause ... 5
3.4 Criticism of the sources ... 6
3.5 Demarcation ... 6
4. Methods ... 7
4.1 Dictionary ... 10
5. Analysis and Result ... 11
5.1 Visborgs Kungsladugård ... 11 5.1.1 Seal ... 11 5.1.2 Other animals ... 13 5.2 Strå ... 13 5.2.1 Seal ... 14 5.2.2 Other animals ... 15 5.3 Gisslause ... 15 5.3.1 Seal ... 16 5.3.2 Other animals ... 17 6. Discussion ... 20 6.1 The seals ... 20
6.1.1 Reasons for the presented distribution ... 23
6.2 Other animals ... 26
6.3 Conclusion... 28
7. Summary ... 29
1
1. Introduction
The early Holocene hunting strategy and seal population has sparked a discussion which has lasted for almost a century. With the finding of the enormous cultural layersof the cave Stora Förvar and the subsequent analysis of the excavated material (Pira 1926) the discussion took form.
With Stora Förvar being the largest and oldest of the sites from the early Holocene on Gotland, this has been an important site for studies of the earliest human activity on Gotland (Lindqvist 1996). Stora Förvar is a cave located on the island of Stora Karlsö outside the southwestern coast of Gotland.
The species of seal present in the Baltic sea during the early Holocene where ringed seal (Phoca hispida) and grey seal (Halichoerus grypus) (Pira 1926; Storå 2001; Schmölcke 2008). The result from the osteological analyses from the oldest layer of Stora Förvar revealed that a majority of the seal present consisted of grey seal with a few elements of ringed seal.
As time progressed the grey seal/ringed seal ratio gradually changed in the cave sequence and ringed seal became more common. This indicated an increase of the hunting of ringed seal at the expense of Grey seal over time. The pattern culminated in the early and Middle Neolithic layers in which there were no grey seals present and the harp seal (Pagophilus groenlandicus) appeared.
Pira (1926) presented two possible hypotheses that could explain the observed pattern. 1) That the hunting of, preferably Grey seal pups on the beaches of Stora Karlsö, had a negative effect on the Grey seal demography over time and
subsequently led to the gradual shift towards ringed seal which requires more sophisticated hunting techniques, or 2) That the changing environment in the Baltic sea basin leading to a warmer climate and higher salinity led to the increase in the Grey seal population. This in turn led to a gradual shift towards the hunting of ringed seal which requires more sophisticated hunting techniques.
When Pira published his work he was not aware of the fact that there is a 2000 year gap in the cultural layerof Stora Förvar during which the cave most likely was flooded by the Litorina Transgression (Lindqvist 1996). When humans returned to the cave after this event the grey seal is all but gone from the archaeological material and the harp seal has taken hold in the Baltic Sea.
The general hypothesis regarding the introduction of the hunting of grey seal in the early Holocene is that this species was relatively easy to catch. It was also the largest animal on the island at that time and from an economic point of view it would make sense to hunt it. Traditionally grey seals have been hunted whilst lying on the shore or on the ice, where the young individuals were targeted and speared / harpooned or clubbed and the adults trying to protect their young has suffered the same fate. The hunting of ringed seal, however, requires significantly more planning and technique, this is most likely the reason for them being hunted to a lesser degree. They are also significantly smaller than the grey seal. Ringed seal has often been
2
hunted close to the shore in breathing holes in the ice. This clearly presents more difficulties and dangers than hunting grey seal on the shore (Lindqvist 1996). The seal was for a long time one of the most important sources of food on Gotland and even in modern times there are a large amount documented locations for seasonal seal hunting. The earliest inhabitants of Gotland were limited in their food resources due to the lack of larger mammals on the Island. Consequently, the hunting of seal played a significant role in the Mesolithic economy. The presence of birds, rabbit and fish in the early Holocene archaeological context demonstrate that there was some economic diversity.
Project
This essay was written as a part of the project “The Pioneer Settlements of Gotland – Early Holocene maritime relations in the Baltic Sea zone” led by Jan Apel (Gotland University) and Jan Storå (Stockholm University). The project address questions regarding the first pioneers of Gotland and their culture. How did the transitioning from a mainly terrestrial hunting strategy to a more maritime one evolve and what effect did this have on the human population. Additionally it will address the question of the change in marine mammal fauna after the introduction of human interaction on the island, with a focus on the grey seal (Halichoerus gryphus) and its much
discussed disappearance from the archaeological material.
1.1 Island fauna
The first pioneers appeared c. 3000 years after mainland Gotland rose up from the sea and the first pioneer phase is dated to the period between approximately 7400- 5500 BC. During this time the island was smaller than today and contained many small lakes and thus presenting a slightly different landscape than today (Figure 1). The late Boreal early Atlantic phase was characterized by a slightly warmer average temperature than today and the large difference that had previously existed between summer and winter began to equal out. A vegetation consisting of hazel, elm, oak, lime, ash and pine could be found on the island. At around 5500 BC the Litorina 1 Transgression caused beach areas to be flooded, which effected the vegetation close to the shore such as pine and elm (Österholm 1989).
3
Fig 1. Gotland at c. 6000 BC (8000 BP cal) with the sites discussed in the text.(Map: Statens geologiska undersöknings SGU).
4
1.2 Earlier research
The study of seal hunting during prehistoric times has been studied by Pira (1926), Lindqvist (1996), Storå (2001), Sten & Wallin (2007) and more. Jan Storå has made an important contribution to the seal hunting studies in the Baltic area (Storå 2001). He has mainly focused on seal hunting during the Neolithic and addresses problems in the osteological material regarding taphonomy and cultural variations. He also presents methods for species identification such as morphological differences and available measurements for species identification. He has published several papers on the issue of seal hunting and species identification (Ericson & Storå 1999; Storå 2000, 2002; Lõugas & Storå 2002). One must also mention Johannes Lepiksaar who has made important work in the field of seal osteology. Though this information has not published it has come to my knowledge through Jan Storå whom has previously had access to his unpublished work as he mentions in Storå (2001).
In Pira (1926) the material from Stora Förvar is presented. In the same reference the cultural layers and subsequently the seal hunting and its evolution as seen in the material is discussed. This was the beginning of the discussion of the earliest seal hunting on Gotland and it has prevailed as a interesting and still unsolved question ever since.
These three sites, Visborgs Kungsladugård, Strå and Gisslause has previously been analysed by Christian Lindqvist whom in connection with the analysis published (Lindqvist 1996) on the subject of the Mesolithic population of Gotland. Further
Munthe & Hansson (1930) presents the first excavation of Gisslause and the material excavated. (Seving 1986) examines the relations and location of Gisslause and Strå as well as Svalings which is another of the earliest dated sites on Gotland. Lindström (2010) takes different approach to the material and the sites and presents
environmental angle to the discussion.
2. Aim and Purpose
The main aim of this essay is to analyze the bone material from the archaeological sites of Strå, Gisslause and Visborgs Kungsladugård in order to answer questions regarding early Holocene hunting strategy on the island of Gotland. The study will focus mainly on the composition of the seal remains and will touch upon topics such as distribution of varies seal species and age. Other distinct changes on the bones such as evidence of human impact will also be of interest and recorded. Further aim concerns the involvement of other animals than seal. The presence of these species will also be of concern whilst forming conclusions with regards to hunting strategy. The study will also address the general discussion regarding the transition into a marine culture and the progress involved.
- Are there any differences in the distribution of different species and age groups?
- Can any conclusions regarding hunting strategies be drawn?
- Is it possible to gain further knowledge about the transition to a marine culture?
5
3. Material
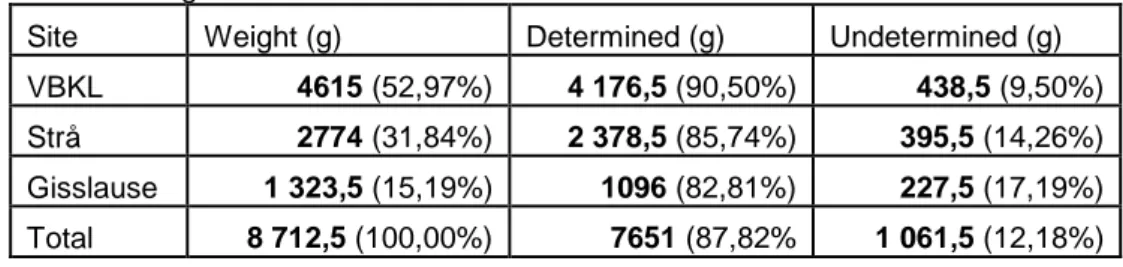
The osteological material from three different sites on the island of Gotland is analysed. These sites are three of the earliest dated sites on Gotland and amongst the few with 14C from the Mesolithic era in Sweden. The majority of the findings consist of seal bones from grey seal and ringed seal. The weight distribution and identification is presented in Table 1.
Table 1. Weight distribution.
Site Weight (g) Determined (g) Undetermined (g) VBKL 4615 (52,97%) 4 176,5 (90,50%) 438,5 (9,50%)
Strå 2774 (31,84%) 2 378,5 (85,74%) 395,5 (14,26%)
Gisslause 1 323,5 (15,19%) 1096 (82,81%) 227,5 (17,19%)
Total 8 712,5 (100,00%) 7651 (87,82% 1 061,5 (12,18%)
3.1 Visborgs Kungsladugård
Visborgs Kungsladugård located 3km southwest of Visby was excavated in 1907 by Oscar Wennersten. Located at around 38m above sea level it has been located slightly over the sea but in close proximity to it. The site is partly covered by gravel from the Litorina 1 Transgression. No human remains have been found on the site, although there is an abundance of evidence of human activity at the site. There are findings of tools such as bone needles, flint-tools, axes, harpoons and other artefacts but the majority of the finds consist of animal bones, mainly seal. Two ceramic
fragments were also found indicating a continuous usage of the site (Lithberg 1914). The material from VBKL was excavated in nine different test pits. The site has been dated to the Early Atlantic period (around 6000-5500BC) (Lindqvist 1996c).
3.2 Strå
Strå is located just west of the town of Fårösund in the northern part of Gotland. The site was previously a gravel pituntil archaeological findings was reported in 1935 and was later excavated in 1936 and 1938 by Mårten Stenberger. During the excavation flint tools and 2 bone items, one harpoon and a fish spear were found. The majority of the material that was recovered consisted of animal bones mainly seal (Seving 1986). The material from Strå was excavated in 22 different test pits. The site has been dated to the Early Atlantic period 6200-5950BC (Lindqvist & Possnert 1997).
3.3 Gisslause
Gisslause located in the northern parts of Gotland, between Visborgs Kungsladugård and Strå. During the Mesolithic the site was located on an esker on a peninsula placing it between the mainland of Gotland and a small island outside in the ocean. Gisslause is the only site to which detailed information about the excavation has been available. Gisslause was excavated in the summer of 1928 by Henrik Munthe and Hans Hansson after having like Strå been a gravel pit for a many years. It was excavated in a half circle with a total of 24 test pits as well as loose findings made at the site without context. Two test pits a few meters away from the main site was also
6
excavated to determine the width and spread of the cultural layer. At the preliminary excavation human bones were located, skull fragments and part of a humerus. During the main excavation however no more human remains were found and the findings consisted only of tools and seal bones (Munthe & Hansson 1930). Unlike the other two sites new excavations has taken place and the site hasn't been destroyed. A minor excavation took place in 1983 where a smaller amount of tools where found together with a few bones (Seving 1986). Unfortunately, other than the artefacts the rest of the material from the 1983 excavation is missing at the present day and has not been found. The summer of 2010 a new excavation took place at Gisslause and a few fragments from bone artefacts and flint blades and cores were recovered. The osteological material from this site consisted of fish, bird, eagle and hare but mainly of seal.
3.4 Criticism of the sources
The main problem with this material is the fact that all of the sites were excavated before modern thorough excavation technique was instated. Visborgs Kungsladugård was excavated over one hundred years ago and a lot has happened since then. These could have resulted in that smaller or more fragmented elements were not found, such as fish vertebrae for example. The sites are not documented sufficiently enough to present the data acquired in a good way, with the exception of Gisslause which was documented quite well by Hansson & Munthe 1928. This causes problems regarding the context of different test pits which can hinder certain questions in the discussion in the present essay. The degree of fragmentation could also cause problems when comparing the sites due to the fact that Visborgs Kungsladugård does have a lower degree of fragmentation than Strå and especially Gisslause with almost 20% of unidentified fragments which is a result of a high fragmentation. This difference in fragmentation is interesting considering that all three sites are located in the same type of ecological environment. They are all covered by the Litorina Transgression and located in a mixture of gravel and sand, this should reasonably put the degree of fragmentation at a relatively equal degree. This
however is not the case concerning Gisslause that has an over 10% higher degree of fragmentation than Visborgs Kungsladugård. The reason for this difference is
unknown, it could be attributed to a more calcareous soil on Visborgs Kungsladugård compared to Gisslause. However, it is possible that this is simply caused by the higher degree of gravel and sand in the soil on Gisslause causing a higher degree of fragmentation.
3.5 Demarcation
This essay will focus on the hunting strategy of seal and the effect on its population during the early Holocene on the island of Gotland. Further, some discussion with regards to marine adaptation on the island and its progress will be addressed. Due to time constraints any detailed analysis and discussion with regards to the presence of remains from humans, birds and sheep / goat will not be presented. Fox and fish will receive a further discussion due to its relevance with regards to the seal population.
7
4. Methods
In answering the aim and purpose of this study, different methods will be used to determine age, sex and species of the seal bones in the material.
Age will mainly be studied through the analysis of the fusing of epiphysis which will be determined to a certain age span (Storå 2001). The following categories will be used to place the bone elements in an age span.
Categories
1. (Sub Adult) Yearlings 1-3yrs (Skeletal age) 2. (Sub Adult) Juveniles 4-5yrs (Skeletal age) 3. (Adult) Young Adults 6yrs (Skeletal age) 4. (Adult) Old Adults 7-8yrs (Skeletal age)
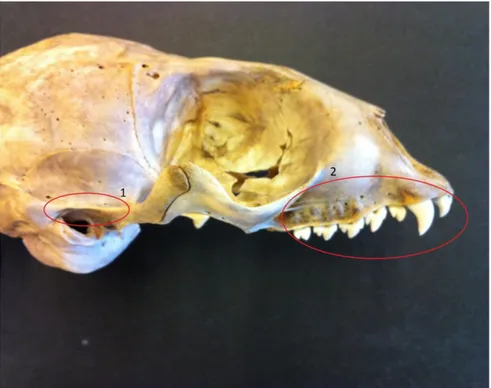
Species will be determined using visual analysis of the temporal bone, teeth and size comparison using fused long bones. The determination of the temporal bone consists of studying the crista below meatus acusticus externa on pars mastoideus where there is a difference in size between the species, where the ringed seal (Phoca hispida) is thin and sharp (Figure 1.1) whilst the grey seal (Halichoerus grypus) is broad and large (Figure 2.1). This makes the temporal bone suitable for species identification and in this essay the identification will mainly focus on this
differentiation.
It is also possible to perform species identification by using size, mainly on long bones. Humerus, radius, ulna, and femur are the best bones long bones for this. These differ significantly in size between grey seal and ringed seal where the grey seal is much larger than the ringed seal. However, in very young individuals the size difference can be hard to determine due to the very small or non identifiable
differences between the species. In this essay species identification using long bones from young individuals has been used with caution due to its difficulties. These
difficulties resulted in that the distribution identified using size was not presented in the results.
Differences between teeth can easily be used for determining seal species. Where the grey seal has large teeth with one point (Figure 2.2) the ringed seal has small teeth with three minor points (Figure 1.2). The canines of ringed seal can be
mistaken for grey seal, however the difference in robustness, size and curvature can be used to determine even those. Due to the fact that each individual seal has a large number of teeth a specific species distribution is difficult to ascertain since five teeth could be from anywhere between one to five individuals.
The problems discussed in chapter 3.4 with regards to excavation technique also poses a problem regarding teeth, as the teeth from the ringed seal is significantly smaller and thus found to a lesser extent, possibly causing a incorrect distribution.
8
Figure 1. Ringed seal. 1. Thin and sharp crista above meatus acusticus. 2. Small multi pointed teeth. Photo by Alexander Sjöstrand.
Figure 2. Grey seal. 1. Broad robust crista above meatus acusticus. 2. Large robust single pointed teeth. Photo by Alexander Sjöstrand.
The osteological bone reference at Gotland University will be used for comparison between species. Associated professor Jan Storå at the Osteoarchaeological
Research Laboratory at Stockholm University has contributed to the visual analysis of species through a personal walkthrough of the differences between species during a visit to Stockholm University by the author in March 2011.
9
For sex determination the existence of the os baculum has been used as evidence for a male individual. Identification of female individuals has been ignored since no accurate published methods for determination exists.
Furthermore, the degree of fragmentation will be based on a visual determination and partly due to the percentage of determined elements. This will be presented in Table 21 showing NISP (Number of identified specimens).
The category Determined requires that the fragment is determined to both species and bone element. A bone element that is determined to a cranial fragment, vertebral fragments or long bones will be regarded as Determined, the specific bone element in these instances will not be required if the species is known. The category
Undetermined will therefore be attributed to fragments which have not been indentified to a specific species or a specific bone element, or both.
The existence of burnt bones will be noted in the results. Burnt bones will be included with the rest of the material when presenting the results. Further information
regarding burnt bones will be presented in the Discussion.
All the material will be weighed according to determined, undetermined and burnt in each test pit. Due to the small amount of non seal bones all these species will be weighed together. All weights will be presented in grams if necessary rounded off 0,5g.
For seal the MNI (Minimum number of individuals) has been calculated using the temporal bone since this bone generally remains well preserved and often found in a representative amount. The MNI of the present material has been calculated using all of the large fragments of pars mastoideus from the left or right side. Since the degree of fragmentation for this element is low this would give a representative image of the MNI.
When presenting the distribution of bone elements, (Table 3, Table 9, Table 15 and Table 22) the categories manus and pedis will exclude phalanges. The phalanges were considered irrelevant for this study and therefore, due to limitation of time, the choice was made not to separate the anterior phalanges from the posterior
phalanges, but instead merely to identify whether it is phalanges 1, 2 or 3. This will result in that the pedis and manus categories will only involve metacarpals /
metatarsals and carpi / tarsi. Additionally, cranial fragments and vertebral fragments will be clumped up as cranial and vertebral fragments and individual bone elements from these categories i.e. cervical vertebrae, will not be distinguished. Mandibula will also be included in the cranial fragments category.
10
4.1 Dictionary
Below, a translation of the Latin names for animals and bone elements which appears in this essay can be seen. The essay will use the Latin terms when
addressing bone elements in the text. The animals will however be referred to their English name, however, together with the Latin name first time it is mentioned. Additionally abbreviations occurring in the essay are presented below.
The sites VBKL - Visborgs Kungsladugård. The species Birds Aves - Birds Fish
Esox lucius - Northern pike Pisces - Fish
Mammals
Capra hircus - Goat
Halichoerus grypus - Grey seal Lepus timidus - Rabbit
Ovis ammon - Sheep
Pagophilus groenlandicus - Harp seal Phoca hispida - Ringed Seal
Vulpes vulpes - Fox
Bone elements
Os temporale - Temporal bone Axis - Second cervical vertebrae Vertebrae - Vertebrae
Scapula - Shoulder blade Humerus - Upper arm bone Ulna - Elbow bone
Radius - Radius Manus - Hand Femur - Thigh bone
Os cruris - Shin bone and calf bone Baculum - Penis bone
Pedis - Foot
Metapod - Metapod Phalanges - Phalanx
11
5. Analysis and Result
5.1 Visborgs Kungsladugård
Fragmentation
The low fragmentation has resulted mostly in large fragments, where the main characteristics remain intact. This resulted in that 50,21% of the total amount of fragments has been determined. The weight of the identified fragments are 90,50% of the total weight (Table 2).
Less than 5% of the total amount of determined fragments consists of slightly burnt bones.
Table 2. Weight distribution and number of fragments. VBKL. Determined Undetermined Total Weight (g) 4176,5 (90,50%) 438,5 (9,50%) 4615 (100%)
Fragments (no) 2441 (50,21%) 2421 (49,79%) 4862 (100%)
Species
Mainly fragments from seal (phoca) have been found from this location. There are, however, some bone elements of other species such as birds (Aves), rabbit (lepus timidus), fox (vulpes vulpes)(Figure 3), sheep / goat (ovis ammon / Capra hircus) and fish (Pisces).
Figure 3. Fox bones. From left to right:
Proximal part of humerus, axis, diafysis of tibia. Photo by Alexander Sjöstrand.
5.1.1 Seal
Bone distribution
The distribution of bones covers the whole body as presented in Table 3, with a large majority of cranium and vertebral fragments. The reason for the large amount of bone fragments from cranium and vertebrae is in all likelihood due to the higher degree of fragmentation of these bones. The fact that the different vertebrates (cervical,
thoracic, lumbar and sacrum) has not been separated, will result in a large number of vertebral fragments. There are however more fragments from the upper long bones (humerus, ulna and radius) than from the lower long bones (femur and cruris) (Table 4) the reason for this difference will be addressed in the Discussion.
12
Table 3. Number of identified fragments from seal. VBKL. Element No. fragments % Cranium 736 36,73% Scapula 25 1,25% Vertebrae 820 40,92% Humerus 99 4,94% Ulna 38 1,90% Radius 50 2,50% Manus 40 2,00% Coxae 37 1,85% Femur 41 2,05% Cruris 48 2,40% Pedis 70 3,49% Total 2004 100,00%
Table 4. Distribution between identified fragments of
humerus, ulna and radius compared to femur and cruris.
VBLK
No. fragments
Upper long bones 187 (67,75%)
Lower long bones 89 (32,25%)
Seal
Identified species are grey seal and ringed seal. For the differentiation of species, the temporal bone was mainly used, which discern a large amount of grey seal.
Altogether 37 grey seal have been indentified and only seven ringed seal (Table 5). Taking the good preservation of the temporal bone into account the distribution between the species is most likely correct, Table 5. This also corresponds with the differentiation of teeth between the species which presents 125 from grey seal and 13 from ringed seal.
Table 5. Number of identified individuals (MNI) from grey seal and ringed seal. VBKL
Grey seal Ringed seal VBKL 37 (84,09%) 7 (15,91%)
Age distribution
The age distribution shows that the majority of the fragments was determined as being Sub-Adult, i.e. that the majority of the population was young, probably in their first few years. Although there are individuals of all ages the older individuals are a minority (Table 6). Where the Sub-Adult and the Adult categories show with certainty fragments from these categories, the other two categories presents a larger age span and cannot with certainty be placed in either the Sub-Adult or the Adult category.
13
Table 6. Number of identified fragments from each age group. VBKL
Sub-Adult
Yearling - Young adult
Juveniles -
Old adult Adult Total VBKL 94 (70,68%) 14 (10,53%) 2 (1,50%) 23 (17,29%) 133 (100%)
MNI
The result from the analysis of MNI resulted in 64 individuals (Table 24). This corresponds to the degree of fragmentation which was low and the general
distribution of bone elements which were numerous and covered the whole body.
5.1.2 Other animals
Due to the small amount of bones from animals other than seal, not much can be revealed. Bone from birds was more commonly found that bones from other animals (Table 7). Bones from rabbit and fox was found in an almost equal amount. Fish and goat / sheep are represented by one or a few fragments each only. The fish
vertebrae that was found has previously been identified as a pike (Esox lucius) (Lithberg 1914).
Table 7. Number of identified fragments from each species. VBKL
Seal Human Fish Rabbit Bird Fox
Sheep / Goat VBKL 2397 (98,2%) 0 (0%) 1 (0,04%) 11 (0,45%) 23 (0,94) 7 (0,33%) 1 (0,04%)
5.2 Strå
Fragmentation
Many of the fragments were 1/3 of the total bone size. This resulted in that 54,30% of the total amount of fragments has been determined. The weight of the identified fragments are 95,74% of the total weight (Table 8).
Less than 6% of the total amount of determined fragments consists of slightly burnt bones.
Table 8. The distribution of weight and fragments. (g) gram, (no) number. Strå.
Determined Undetermined Total
Weight (g) 2378,5 (85,74%) 395,5 (14,26%) 2774 (100%)
Fragments (no) 1478 (54,30%) 1244 (45,70%) 2722 (100%)
Dating
Strå has been dated to the Early Atlantic period 6200-5950BC (Lindqvist & Possnert 1997)
Species
The material from Strå mainly consists of seal (grey seal and ringed seal). Only few bones from animals other than seal were identified, such as rabbit, birds and fish (Table 13).
14
5.2.1 Seal
Bone distribution
The distribution of bones covers the whole body as presented in Table 9. There are however more fragments from the upper long bones (humerus, ulna and radius) than from the lower long bones (femur and cruris) (Table 10).
Table 9. Number of identified fragments from seal. Strå Element No. fragments % Cranium 393 48,16% Scapula 12 1,47% Vertebrae 100 12,25% Humerus 38 4,66% Ulna 27 3,31% Radius 37 4,53% Manus 29 3,55% Coxae 19 2,33% Femur 25 3,06% Cruris 44 5,39% Pedis 92 11,27% Total 816 100,00%
Table 10. Distribution between identified fragments of humerus, ulna and radius compared to
femur and cruris. Strå
No. identified fragments
Upper long bones 102 (59,65%)
Lower long bones 69 (40,35%)
Seal
Due to the low amount of temporal bones and the difficulties in differentiating long bones a low amount of fragments has been identified from seal. Five grey seal and six ringed seal have been identified (Table 11). This does not correspond with the distribution of teeth where 96 teeth from grey seal and seven teeth from ringed seal have been identified.
Table 11. Number of identified individuals (MNI) from grey seal and ringed seal. Strå
Species Grey seal Ringed seal Strå 5 (45,45%) 6 (54,55%)
15
Age distribution
The age distribution demonstrates a slight majority of individuals in the Adult category, indicating a older rather than younger population (Table 12). Where the Sub-Adult and the Adult categories show with certainty fragments from these categories, the other two categories presents a larger age span and cannot with certainty be placed in either the Sub-Adult or the Adult category.
Table 12. Number of identified fragments from each age group. Strå Sub Adult
Yearling - Young adult
Juveniles -
Old adult Adult Total Strå 23 (28,05%) 6 (7,32%) 19 (23,17%) 34 (41,46%) 82 (100%)
MNI
The bones from Strå represent a minimum number of 11 individuals (MNI) (Table 24).
5.2.2 Other animals
The majority of bones from other animals than seal are from birds, represented by 14 fragments. Together with four fish vertebrae and two metapods from rabbit these animals do not allow any further analysis (Table 13).
Table 13. Number of identified fragments from each species. Strå
Seal Human Fish Rabbit Bird Fox
Sheep / Goat Strå 1458 (98,65%) 0 (0%) 4 (0,27%) 2 (0,14%) 14 (0,95%) 0 (0%) 0 (0%)
5.3 Gisslause
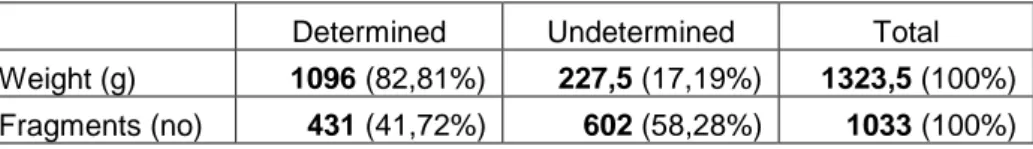
Fragmentation
The degree of fragmentation resulted in that 41,72% of all fragments were
determined, these fragments represented 82,81% of the total weight of the material (Table 14).
Less than 1% of the total amount of determined fragments consists of slightly burnt bones.
Table 14. The distribution of weight and fragments. (g) gram, (no) number). Gisslause.
Determined Undetermined Total
Weight (g) 1096 (82,81%) 227,5 (17,19%) 1323,5 (100%)
Fragments (no) 431 (41,72%) 602 (58,28%) 1033 (100%)
Species
Human remains has been identified, however these consist of only few fragmented bone elements: fragments from the skull and from the diaphysis of a humerus. The identified animals are seal, rabbit, bird and fish.
16
5.3.1 Seal
Bone distribution
The distribution of bones covers the whole body as presented in Table 15, there are however more fragments from the upper long bones (humerus, ulna and radius) than from the lower long bones (femur and cruris) (Table 16).
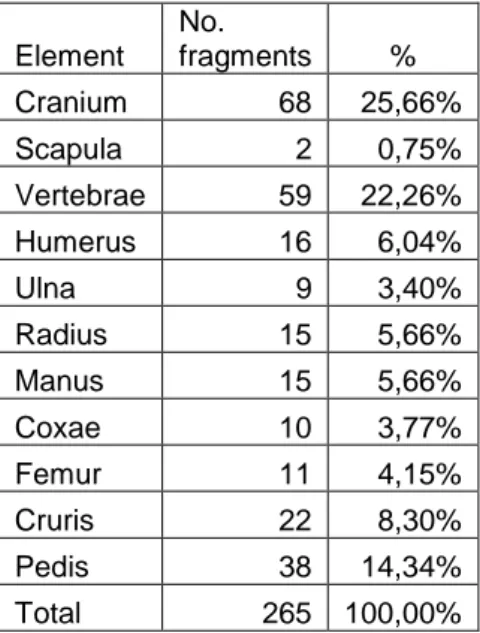
Table 15. Number of identified fragments from seal. Gisslause.
Element No. fragments % Cranium 68 25,66% Scapula 2 0,75% Vertebrae 59 22,26% Humerus 16 6,04% Ulna 9 3,40% Radius 15 5,66% Manus 15 5,66% Coxae 10 3,77% Femur 11 4,15% Cruris 22 8,30% Pedis 38 14,34% Total 265 100,00%
Table 16. Distribution between identified fragments of humerus, ulna and radius compared to femur and cruris. Gisslause.
No. Identified fragments Upper long bones 40 (54,79%)
Lower long bones 33 (45,21%)
Seal
Species determination was problematic due to the small amount of bone material and the high degree of fragmentation. With only two individuals identifiable with the
temporal bone from grey seal and three fragments from ringed seal (Table 17) a true distribution between the species at Gisslause is hard to ascertain. However, from the material available no other distribution can be presented.
Table 17. Number of identified individuals (MNI) from grey seal and ringed seal. Gisslause
Species Grey seal Ringed seal Gisslause 2 (40,00%) 3 (60,00%)
17
Age distribution
The age distribution of fragments from seal on Gisslause show a quite equal distribution where there is no major difference between the Sub-Adult and Adult categories (Table 18). Where the Sub-Adult and the Adult categories show with certainty fragments from these categories, the other two categories presents a larger age span and cannot with certainty be placed in either the Sub-Adult or the Adult category. However, due to the small size of the material from Gisslause this might not be representative of the site as a whole.
Table 18. Number of identified fragments from each age group.
Sub-Adult
Yearling - Young adult
Juveniles -
Old adult Adult Total Gisslause 16 (36,36%) 4 (9,09%) 9 (20,45%) 15 (34,09%) 44 (100%)
MNI
The MNI analysis shows a minimum number of seven individuals (Table 24). But, according to the small sample size it is difficult to acquire a representative MNI.
5.3.2 Other animals
Only few fragments have been identified from other animals than seal. Identified fragments are from fish, rabbit and bird, but the low number of bones from each species makes it impossible to ascertain further information (Table 19).
Human remains have also been identified. These fragments consist of 3.71 % of the total number of bone fragments from Gisslause (Table 19). All fragments, except one which is from the diaphysis from a humerus, derive from the cranium. The identified human cranial bones consist of a slightly fragmented frontal bone, and three
fragments of the parietal bone.
Table 19. Number of identified fragments from each species.
Seal Human Fish Rabbit Bird Fox
Sheep / Goat Gisslause 396 (91.88%) 16 (3,71%) 9 (2,09%) 5 (1,16%) 5 (1,16%) 0 (0%) 0 (0%)
5.4 Summary, results
Strå, Visborgs Kungsladugård and Gisslause present interesting differences. Table 20 presents the distribution of species and number of fragments for each species. A clear majority of seal fragments can been seen, followed by birds, rabbit and fish. It can also be seen that VBKL is the only site where fox and sheep / goat are present and likewise Gisslause is the only site where human remains have been located. Table 21 shows the ratio of determined fragments. Where both Strå and VBKL present over 50% identification rate with 54,30% (Strå) 50,21% (VBKL) a difference can be seen for Gisslause where only 41,72% of the total amount of fragments has been identified.
18
Table 20. Distribution of species. Number of identified fragments.
Seal Human Fish Rabbit Bird Fox
Sheep / Goat VBKL 2397 (98,2%) 0 (0%) 1 (0,04%) 11 (0,45%) 23 (0,94) 7 (0,33%) 1 (0,04%) Strå 1458 (98,65%) 0 (0%) 4 (0,27%) 2 (0,14%) 14 (0,95%) 0 (0%) 0 (0%) Gisslause 396 (91,88%) 16 (3,71%) 9 (2,09%) 5 (1,16%) 5 (1,16%) 0 (0%) 0 (0%) Total 4251 (97,75%) 16 (0,37%) 11 (0,32%) 18 (0,41%) 42 (0,97%) 7 (0,16%) 1 (0,02%)
Table 21. Number of identified and unidentified fragments. Determined Undetermined
VBKL 2441 (50,21%) 2421 (49,79%)
Strå 1478 (54,30%) 1244 (45,70%)
Gisslause 431 (41,72%) 602 (58,28%)
Total 4350 (50,48%) 4267 (49,52%)
An interesting difference can be seen between VBKL opposed to Strå and Gisslause. As seen clearly in Figure 3 there is a peak in fragments from humerus on VBKL where the rest of the distribution of identified bones remains equal between the sites. The Strå and Gisslause materials show a distribution where there are only minor differences between the bone elements with the exception of pedis which presents a peak at all three sites. Most likely due to the large amount of pedis fragments present with each individual, additionally the slightly larger size than the manus bones makes them easier to find during a excavation. (Figure 3, Table 22).
There is a difference between the occurrence of upper and lower long bones between the sites, where of VBKL upper long bones was found in a much higher degree. However, this difference is minor on Gisslause but instead more evident on VBKL and Strå (Table 23).
19
Table 22. Number of identified bone fragments from seal.
VBKL Strå Gisslause Cranium 736 (36,73%) 393 (48,16%) 68 (25,66%) Scapula 25 (1,25%) 12 (1,47%) 2 (0,75%) Vertebrae 820 (40,92%) 100 (12,25%) 59 (22,26%) Humerus 99 (4,94%) 38 (4,66%) 16 (6,04%) Ulna 38 (1,90%) 27 (3,31%) 9 (3,40%) Radius 50 (2,50%) 37 (4,53%) 15 (5,66%) Manus 40 (2,00%) 29 (3,55%) 15 (5,66%) Coxae 37 (1,85%) 19 (2,33%) 10 (3,77%) Femur 41 (2,05%) 25 (3,06%) 11 (4,15%) Cruris 48 (2,40%) 44 (5,39%) 22 (8,30%) Pedis 70 (3,49%) 92 (11,27%) 38 (14,34%) Total 2004 (100%) 816 (100%) 265 (100%)
Table 23. Upper and lower long bone distribution.
VBKL Strå Gisslause
Upper long bones 187 (67,75%) 102 (59,65%) 40 (54,79%)
Lower long bones 89 (32,25%) 69 (40,35%) 33 (45,21%)
Due to the largest material being from VBKL a large MNI has been acquired from that site whereas Strå and Gisslause, with a smaller material and a higher degree of fragmentation, show a lower MNI (Table 24).
There is a difference in the distribution of species between the sites, where VBKL demonstrate a majority of grey seal. In contrast, Strå and Gisslause, although with fewer identified fragments, demonstrates a more equal distribution (Table 25).
Table 24. Distribution of MNI for seal.
MNI
VBKL 64
Strå 11
Gisslause 7
Table 25. Distribution of seal species. Number of identified individuals. (MNI)
Grey seal Ringed seal VBKL 37 (84,09%) 7 (15,91%)
Strå 5 (45,45%) 6 (54,55%)
Gisslause 2 (40,00%) 3 (60,00%)
As with the distribution of species, the distribution of age shows a difference between VBKL opposed to Strå and Gisslause. VBKL shows a majority of fragments from the Sub-Adult category whereas Strå and Gisslause shows a more balanced distribution, with a slight majority of fragments from the Adult category on Strå and a equal
distribution on Gisslause (Figure 4, Table 26). Where the Sub-Adult and the Adult categories show with certainty fragments from these categories, the other two categories presents a larger age span and cannot with certainty be placed in either the Sub-Adult or the Adult category.
20
Figure 4. Age distribution. Number of bone fragments from each category
Table 26. Distribution of age. Number of bone fragments from each age category.
Sub-Adult
Yearling - Young adult
Juveniles -
Old adult Adult Total VBKL 94 (70,68%) 14 (10,53%) 2 (1,50%) 23 (17,29%) 133 (100%)
Strå 23 (28,05%) 6 (7,32%) 19 (23,17%) 34 (41,46%) 82 (100%)
Gisslause 16 (36,36%) 4 (9,09%) 9 (20,45%) 15 (34,09%) 44 (100%)
6. Discussion
6.1 The seals
Due to the fact that most of the osteological material from Gotland consists of seal this has been the major theme of discussion when presenting hunting strategies for the early Holocene population. There has been a discussion regarding why the seal population changed over time, mainly grey seal. One hypothesis states that the grey seal population was negatively affected by the hunting of mainly the young
individuals, that were clubbed, speared or harpooned at the shore during their breeding period (Pira 1926).The disappearance of grey seal could also be attributed to the changing climate and increased salt level and colder climate. This would have changed the habitat of the grey seal making it viable for a migration of harp seal into the Baltic. The effect of this could be reflected in the gradual change from grey seal to harp seal in faunal remains of the early sites (Lindqvist 1996c).
For whatever the reasons the change did occur it apparently forced humans to evolve their hunting strategies and technique. It pushed them to hunt ringed seal with
harpoons because of the fact that the ringed seal probably were hunted on the ice, since they require breathing holes in the ice during the winter (Lindqvist 1996).
21
Visborgs Kungsladugård
VBKL presents a rather young population reasonably in their first year, with a large majority of grey seal (37 grey seal versus seven ringed seal). This distribution would place this in context with hunting strategies during the Bronze Age in the Baltic, which would indicate a hunting season during the late winter / early spring (Lõugas & Storå 2002) corresponding to the breeding period for grey seal in the Baltic Sea today. Additionally the large difference between upper and lower long bones can most likely be attributed to natural fragmentation because of the fact that humerus, radius, ulna are smaller and more fragile than femur and cruris, taking into account that cruris is two bones tibia and fibula combined which also makes the bone more robust in the proximal end, resulting in a lower degree of fragmentation.
Strå
The material from Strå consists of a smaller sample than VBKL, despite this an interesting difference has can be seen. Strå displays an almost equal distribution of the species (five grey seal versus six ringed seal) and the age distribution is also almost equal, with a slight majority of older seals. This matches the result by Storå (2002) where he points out that the youngest and the older individuals in the ringed seal population would be affected the most by hunting, provided that the hunt did occur during breeding season.
Gisslause
The composition of the Gisslause material is similar to the one found on Strå where the age and species distribution is equal. With only a few identified grey and ringed seal (two grey seal, three ringed seal) and a similar age distribution.
Criticism of sources
Before any conclusions are drawn, further possibilities to the age and species distribution must be explored. The species distribution has been performed on the temporal bone (os temporale) which is part of the cranium. As mentioned in Storå (2001) cultural effects such as a culinary preference or transportation of the bones / individual could result in a non-representative distribution in the archaeological material.
To present an example, take VBKL where the majority of identified seal bones belong to grey seal. These were identified using cranial bones, if these bodies has been transported it is not unlikely that they were dismembered before transportation to ease the transportation process. With this proposal in mind imagine ringed seal being transported from the north-eastern coast (Strå, Gisslause) to the west coast where VBKL is located. Such a procedure could result in an absence of cranial bones from ringed seal since these has been severed from the body before transfer.
An interesting aspect of the results is that there are more grey seal teeth compared to ringed seal teeth on Strå. This contradicts the species identification which presents only five grey seals an six ringed using the temporal bone. This presents an
interesting dilemma, is may be possible that the species distribution on Strå is more similar to VBKL where the grey seal is clearly dominant to the ringed seal but
22
because of some unknown element the distribution of the temporal bones does not show the same distribution as the teeth. As presented in chapter 4 this could be due to the excavation technique used on the two excavation sites. Neither of these sites was thoroughly excavated. This may have resulted in a tendency in which smaller teeth of the ringed seal were not recovered during the excavation as opposed to the slightly larger more robust teeth from the grey seal. Before any premature
conclusions are drawn, the fact that both the temporal bone and the teeth derive from the cranium has to be acknowledged. This does strengthen the probability of the species distribution presented by the analysis of the temporal bone since this is a robust bone element which should be found in an equal amount between the species due to the insignificant size difference between them.
An important fact is that on VBKL two pieces of ceramics was found, indicating a possible mixture of cultural layers. Since VBKL unlike Strå and Gisslause is not fully covered by the Litorina Transgression this could have caused contextual problems. However, when considering this problem the fact that only two pieces of ceramics where recovered which would not indicate a complete mixture. Additionally the fact that a sheep / goat bone has been identified could strengthen the theory of a mixed cultural layer, unless this single bone element was excavated in a test pit where no other finds where discovered indicating an entirely different context. However, the results strengthen that there is little or no mixture since there should be a larger amount of ringed seal amongst the material if it was dated to the Neolithic or later, one would also expect more animals not found in the Mesolithic materials such as large sea fish and pigs.
One must also keep in mind that the smaller samples from Strå and Gisslause could result in a misreading of the results, the distribution that is found on both sites might not be representative of the material originally deposited on the sites. Especially the species distribution can be questioned due to the small amount of identified
fragments. These are problems that will have to be kept in mind when conclusions are presented.
What also has to be noted is that further knowledge with regards to the determination of species, i.e. distribution of species, could have been obtained if the time permitted this when visiting Jan Storå at Stockholm University. The surplus of knowledge concerning the identification of long bones could have presented a different result as well as strengthened the present distribution.
It also has to be said that the temporal bone should present a correct distribution to a larger degree than the long bones. Due to the larger size of grey seal long bones compared to ringed seal long bones, this could result in a skew distribution due to excavation technique and the preservation of the bones. Where the temporal bone is robust and equal in size between the species this element should be found to a equal extent.
Additionally it has to be clarified that the test pits has been clumped up and all of the material from each site has been treated as one context and no differentiation
between the test pits has been made. Whilst this should not have any impact on the results it has to be mentioned.
23
6.1.1 Reasons for the presented distribution
The apparent difference between these sites can be seen in the age and species distribution where VBKL presents a majority of young grey seals whilst Strå and Gisslause presents a rather equal distribution looking at both age and species. These differences present themselves despite the relatively similar dating which presents an interesting question, why are these differences present? To answer this three
different theories has been proposed, these will be presented below.
If there were different contemporary populations hunting seal on the island, the young grey seals identified on VBKL would be indicative of a slightly less advanced hunting strategy where the population has simply clubbed, speared or harpooned the seals on the shore and brought them home. When compared to Strå and Gisslause, however, where the age distribution is more equal or even slightly in favour of older individuals whilst the species distribution also seems more equal in comparison. This distribution could give a possible indication of a more advanced hunting strategy where the hunt has been taken place further out to sea using harpoons to hunt down older ringed seal through their breathing holes.
Would this then be indicative of separate populations with a need for different hunting strategy due to their location (VBKL, Midwestern Gotland. Strå, Gisslause, North-eastern Gotland)?
This is of course a possible scenario, however, I believe that it is unlikely due to the small size of the island and the short distances between the sites. It is less than half a day’s walk between the North-eastern sites of Strå and Gisslause to the
Midwestern site of VBKL.
Another possible scenario is that the pattern reflects seasonal hunting strategies by the same population? The Middle Neolithic seal hunting on Åland occurred during late winter / early spring and mainly consisting of the hunting of ringed and harp seal. Bearing in mind that the ringed seals breed during this time and that the juveniles in the population are separated from the rest of the age groups at this time the yearlings and adults would have suffered the most (Storå 2002).Bronze Age seal hunting mainly occurred during late winter / early spring and as far as early summer. The seals analysed from Bronze-Age sites mainly consist of grey seal and harp seal (Lõugas & Storå 2002) and this indicate that the hunting of grey and ringed seal coincided season wise. This is however well after the disappearance and return of the grey seal in the Baltic, which might have affected the breeding pattern and overall behaviour of the grey seal. This could therefore show a different breeding pattern than would have been present during the early Holocene.
The grey seals age distribution on VBKL does match the Bronze Age hunting season presented by Lõugas & Storå (2002) indicating that the seal hunt did occur during the grey seal breeding period i.e. during late winter / early spring. Additionally it is
possible that the grey seal was hunted during the summer on VBKL, the age
distribution in the present does not prohibit this. If grey seals where hunted during the summer this would still present a large majority of young individuals, which is the case in the VBKL material. This is also true for the seals identified from Strå and Gisslause which also match the data presented by Storå (2002), where younger and older individuals are equally represented in the materials. However, due to the
24
fragmentation of the material the actual distribution is hard to ascertain and because the juveniles should not be present due to the fact that they are apart from the rest of the population during the breeding season i.e. when the hunt should have occurred. This makes it hard to prove this hunting strategy with certainty.
A seasonal hunt is possible, although it is hard to prove due to the uncertainty of the Strå and Gisslause materials. However, the idea of a seasonal hunting strategy should not be dismissed. If the grey seals was in fact hunted during the summer this would present a strong argument for a seasonal hunt. Seasonal hunting of seals is not unheard of and Storå (2002) presents data pointing to a ringed seal hunt occurring in late winter / early spring whilst the hunting of harp seal occurred later during the year approximately late spring / early winter. If this was the case with the early Holocene gotlandic population, the Mesolithic population on Gotland would have access to a rich fauna of seals year round.
A combination of both?
Could it simply be that one population out of necessity and understanding of the fragility of the seal population had different hunting sites which they used
periodically? The slow reproduction and high pressure on the seal demography due to hunting of young individuals would have had a high potential for changing the population negatively. If this was understood by the Mesolithic population, would it not be reasonable to assume that they would migrate to other parts of the island to ascertain a stable fauna, and then return to their former site? Such a settlement pattern was most likely based on some seasonality where they moved with regular intervals. Their main subsistence source would most likely have been the grey seal due to its size and the fact that they was easier to hunt whereas the ringed seal became a complement when the grey seal population decreased.
This actually seems like the most probable hypothesis; a partial seasonal hunt would be expected due to the lack of other sources of food to sustain a population on the island. Storå (2002) argue that a seasonal seal hunt has been present in prehistoric times. How this seasonal hunt originated and to what extent it was in use is at the present time impossible to determine. The study presented in this essay indicates that the possibility of this is very high and cannot be dismissed without further evidence to the contrary.
Marine adaptation
The discussion on marine adaptation and the transition from terrestrial hunt to a marine hunt does present some interesting topics.
The three analyzed sites do present a relatively new ecological adaptation, where the inhabitants on the island have previously mainly been dependant on the fishing of fresh water fish and the hunting of terrestrial animals. Accordingly, the specific environment of Gotland forced the population to quickly adapt to a new fauna with a need for new tools and hunting strategies.
(Bjerck 2009) divides marine adaptation into three categories or relations to a marine environment as Bjerck calls it. These three are lacustrine, littoral and marine
25
and such which are used for transportation, fishing and movement control for pray such as deer etc. This is the first stage of marine relations without any direct interaction with the open ocean. The next stage is referred to as littoral, and is defined by the use of beaches etc, a increase in marine usage has occurred and gathering activities such as gathering shells, seaweed and items related and found in and near ocean areas has large part in the day to day lives of the population. Fishing at the shore is also included in this category as well as hunting seals close to and on the shore, which does include hunting on the ice through breathing holes, or
scavenging for washed up animals at the beach. The last stage is a full marine adaptation where the open sea is available, this requires a good knowledge of the open ocean, weather conditions and boat building. The ocean can now be used for fishing and hunting on open sea as well as travelling for longer distances in more open seas (Bjerck 2009).
The fact that the majority of the identified bone material derives from marine mammals i.e. seal, a partial marine adaptation can be observed on the early
Holocene sites of Gotland. This would correspond to Bjercks littoral relation, where the population has not yet began to open up the full potential of a marine culture but has begun using marine resources. With most of the hunting based on hunting seals on the shore or on the ice close to the shore are the first steps in an adaptation to a full marine relation. One could imagine that their seafaring skills would have been on a not to primitive level, considering the migration to the island. Although it has been proven that it is possible to paddle from the mainland to the island in a day
(Österholm 1989).
According to this scenario, a full marine adaptation on Gotland did not fully occur until the Neolithic with the Pitted Ware Culture, where the open sea hunting of harp seal as well as open sea fishing were practised This focus on seal hunt can be seen as the first step in the direction of the later Pitted Ware Culture and their knowledge of open sea hunting and travelling.
Colten and Arnold (1998) classify animals in two categories, r-selected and K-selected, where r-selected are fast reproducing, fast growing and large in numbers, (for example fish and shellfish) while K-selected animals have a slower reproduction rate, grow slower and are fewer in numbers (for examples mammals and birds). R-selected animals can support a large population due to their constant increase in population despite pressure due to hunting. K-selected animals can only support a small population over a longer period since a high hunting pressure could have a devastating impact on a K-selected population, especially if the young individuals of a population is targeted, as has been traditionally been the case whilst hunting grey seal and ringed seal.
26
6.2 Other animals
The absence of fish
The almost complete absence of fish in the material from the sites raises an interesting question. Has the nutrition intake been entirely based on seals and
vegetables despite the presence of a probable large amount of freshwater fish within reach from every location on the island? An answer to this question would present a relevant addition to the discussion of this essay, however, the virtual absence of fish on the sites could be attributed to the excavation techniques or to the poor
preservation of the fragile vertebrae common in archaeological materials. All three sites were first excavated before the middle of the century, before the relevance of proper and detailed excavation after even the smallest fragments. This could have had a huge impact on the finding of fish and even smaller birds which might not have been as well preserved or large in fragment size as the seal bones amongst the material.
One could argue that freshwater fish would be a good complement to their heavy diet of meat from seal. Colten & Arnold (1998) discuss the relationship between fishing and marine mammal hunting, and conclude that the presence of fish does increase when marine mammal remains decrease. This could explain the lack of fish remains. They further argue that marine mammals can only support a relatively small
population due to their reproductive time compared to fish, for example. A seal population would quickly diminish if a large population would depend on it for
sustenance, whilst a fish population would be able to sustain a large population due to their much faster reproduction rate.
The change in the grey seal population might thus be attributed to an increase in population, pushing the grey seal population to its limit, just as Pira (1926) discussed. This would also have triggered the increase of the fishing in lakes and the ocean that can be observed on Neolithic sites such as Ajvide where fish does present a large compliment to seal and pig in the archaeological material (Burenhult 1999).
Acknowledging that this is still early on in the history of the island, the population was in all likelihood not large. This would support that fish was not as important during this period as it later would become after the change that occurred with the Litorina
Transgression and the subsequent climate change and migration of new species to the island, such as pig and fox. Although fox being present at VBKL that population was in its first stage and probably small in size during this time.
The lack of larger ocean fish such as cod could be attributed to the argument discussed by Bjerck (2009) where it is put fourth that it is not until the last stage in marine development that open ocean fishing would occur. A stage seemingly not reached by the early Holocene population of Gotland. Due to the large size of the vertebrae from fish such as cod, these should not have been effected by the excavation technique and can thus been seen as not present.
It must also be presented that during the excavation of Gisslause during the summer of 2010, I personally observed a much larger amount of fish fragments than
presented by these results, this is most likely due to the much more thorough
excavation technique applied in 2010 where we used water silting compared to older excavations where it is unclear if they have even silted the soil. This would in all
27
likelihood change these results, presenting a much larger amount of fish. However from the present material this distribution cannot be proven.
The presence of fox
The fox, the largest mammal on the island for a long time, is only present at VBKL. Is this simply a coincidence or can it be explained by a slightly separate dating? Does this site have a more complex chronology? VBKL is located on the other side of the island, could this indicate that the foxes habitat was limited to this area and was absent in the northern regions. As VBKL does present the latest dating, this may explain the presence of fox on VBKL. These are however the earliest findings of fox on Gotland. The fox population was in all likelihood small at this time, which could also limit them to a certain part of the island from where they spread later on.
Sheep / Goat
The sheep / goat bone element found on VBKL is an odd finding and indicate that the site is mixed. It is important to remember that VBKL is only partially covered by the Litorina Transgression layers and that parts of the site could contain chronologically mixed materials.
Human remains
Gisslause is the only one of these three sites with human remains, the reason for this is unknown. The possibility of a burial site further inland is not impossible or whether this is simply due to an unintended mixture of animal and human remains buried in the area for a unknown reason. Never the less these human bones found on Gisslause are known as some of the oldest human remains on Gotland.
Unfortunately the human remains does not have any context, which makes further analysis difficult.
Burnt bones
The presence of burnt bones, most of which are slightly burnt in the material does not indicate something relevant to the discussion, due to the fact that the bones are only slightly burnt this is merely an indication that the bones have been held over a fire, probably whilst cooking the meat. Whilst some are more burnt than others if only by a slight degree, these can be seen as having been laying in the fire in addition to being burnt whilst cooking the meat.
28
6.3 Conclusion
- Are there any differences in the distribution of different species and age
groups?
It would be overstepping the results to draw any definitive conclusions from the present study. There are, however, some interesting results. The difference between the western located site of VBKL and the north-eastern sites of Strå and Gisslause presents an interesting dilemma. Where VBKL shows a young population of mainly grey seals Strå and Gisslause shows a more balanced distribution of age and species with Strå presenting even a slight majority of older individuals.
- Can any conclusions regarding hunting strategies be drawn?
The differences between the sites may reflect different groups living on different locations on the island. However, this seems highly unlikely due to the small size of the island and the relatively short distances between the locations. This is no more than a few hours walk which makes is highly unlikely for two groups with different hunting patterns to coexist on the same island at the same time period.
The most likely theory that can be drawn from this material is a seasonal hunt, or at least partially seasonal. This could explain the difference. As presented, grey seals have traditionally been hunted on the shore during their breeding period while the ringed seal has been hunted through breeding holes out on the ice. This represents two different strategies that could have been used by the same population due to high pressure on one population they would resort to another population in order for the other one to recover. Further this would most likely point to difference in seal populations around the island, one major grey seal population at the Midwest-coast and a more spread seal population located further up the north-eastern coast.
- Is it possible to gain further knowledge about the transition to a marine
culture?
Seeing that the majority of the material consists of grey and ringed seal, there is no evidence to suggest open ocean hunt. The lack of large deep sea dwelling fish also supports this claim. The early Holocene population can be seen in a transitioning phase between a previous terrestrial sustenance to a marine one, where they have not yet taken the step out into the ocean to explore its full potential. A step that was not taken until a few thousand years later with the Pitted Ware Culture.
29
7. Summary
The material consists of three sites, Visborgs Kungsladugård, Strå and Gisslause. Visborgs Kungsladugård presented a majority of young grey seals, whilst on the other two sites, Strå and Gisslause there is a more equal distribution between grey and ringed seal. The difference is interesting due to the fact that Visborgs
Kungsladugård is located on the Midwestern coast of Gotland whilst Strå and Gisslause are located on the north-eastern coast. According to the results this suggests a different hunting strategy on the western coast contrary to the north-eastern. This indicates a partial seasonal hunt which has been known to occur later on during prehistoric times.
The focus on seal hunting indicates the littoral stage in the marine adaptation, where too population mainly focus on hunting and gathering close to the ocean shore and in fresh water bodies. Though they had not yet begun to exploit the full potential of the oceans resources.