Boplatsval hos solitära getingar, bin och rovsteklar
Examensarbete i biologi, inriktning ekologi, nivå D magisterav Torkel Fischer, januari 2009
Summary
This study deals on the issue of nest-choice among solitary trap-nesting hymenopteras. Trap-nests were constructed and made availiable as nesting substrate. The result showed that the hymenopteras had a tendency to nest in aggregations, whereas there was no preferens for central nesting positions. The hymenopterans also showed a tendency to nest on left part of the trap-nest surface. Aggregation could be a way to avoid parasitation and predation in the nesting area. However, if this was the cause for aggregation, it would be unexpected that no central tendency was indicated. It is hard to suggest a reason why hymenopterans should prefer to nest on the left part of the available substrate.
Sammanfattning
Denna undersökning gjordes för att studera solitära steklars boplatsval speciellt med avseende på spatiala mönster. Aggregat av lämpliga bohål (bibatterier) tillverkades och gjordes tillgängliga förfrilevande steklar. Resultatet visade att steklar tenderar att anlägga bon i närheten av andra stekelbon. Ingen tendens fanns att anlägga bon i centralt belägna positioner. Steklarna verkar också föredra att anlägga bon till vänster om en tänkt mittllinje på bibatterierna. Aggregation kan vara ett sätt att undgå parasitering och predation vid boplatsen. Om detta orsakade aggregationstendensen var det dock något oväntat att inte också en central tendens kunde påvisas. Att steklarna föredrog bohål i bibatteriernas vänstra del är svårt att förklara.
Inledning
Steklar (ordning Hymenoptera) utför för människan nyttigt arbete genom skadedjursbekämpning och pollinering (Banaszak 1992, Cane & Tepedino 2001, Williams 2002, Larsson 2006). Framför allt pollineras de vilda växterna. Utan dessa växter finns inga vilda steklar, flugor och fjärilar vilket medför att människans grödor överhuvudtaget ej bli pollinerade annat än av tambiet. Av alla kända växter är 75% är beroende av att insekter överför pollen till pistillens märke (Kullenberg 1949). Steklarna och andra pollinerande insekter hjälper alltså till med att hålla artrikedomen på en hög nivå avseende mängd, antal och utbredning, samt att de själva ingår i artrikedomen i naturen. Granvik (1928) formulerade sin syn på naturen (här något förkortat), att den - med alla sina levande väsen - också är till för sin egen skull och icke, som vi mången gång mena, endast för vår skull. Steklar har genom sin pollinering och insektspredation en stor plats i ekosystemen och de utgör födokälla för fåglar och nedbrytare och är nisch-skapare för parasiter, de får därmed en placering medelhögt i näringskedjan.
Steklarna har många olika levnadssätt. En del solitära bin, getingar, rovsteklar och parasitsteklar från underordningarna Apocrita och Parasitica bygger bon i ihåliga kammare ovan mark. De anlägger bon i trägångar efter skalbaggar, tomma snäckskal (Gess & Gess 1999), och i växtstjälkar, som de i vissa fall kan gräva ut själva (Pettersson 2003). Det kan uppskattas att ca 150 arter i Europa (inklusive parasitsteklar) kan anlägga sina bokammare i av människan tillverkade boplatser, i form av borrade hål i trä, buntar av vassrör eller dylikt. Dessa artificiella hål kallas bibatteri och kan användas till
inventeringar, och, framför allt, till att rädda hotade restpopulationer (Gaston et al. 2005). Det finns en fauna motsvarande bibatteristeklarna som lever i marken eller anlägger fribyggen. Kring ovan nämnda steklar är en stor mängd parasitstekelarter knutna.
Steklar som bebygger bibatterier förekommer huvudsakligen på öppna ytor med gamla ängar och betesmarker där det finns en rik flora och insektsfauna nödvändig för steklarnas födosök. Därtill måste det i habitatet finnas solexponerad torr död ved och en ständig tillförsel av ny död ved för att skapa nya förutsättningar allteftersom veden förbrukas. Steklarna utnyttjar håligheter i veden som
uppkommer efter larvgångar av skalbaggar (xylofager) och ihåliga horisontella och vertikala
växtstjälkar (personlig observation). Artspecifika krav kan vara avgörande bland annat att vissa växter måste finnas i habitatet när steklarna är oligolektiska dvs. beroende av en specifikt växtgrupp.
Utbredningarna är ofta fragmenterade och det kan visas genom att jämföra inventeringar från olika områden (Quaranta et al. 2004).
Ordningen steklar är mycket viktig vid arbetet med biologisk restaurering (Loyola & Martines 2006). Det finns många hot mot stekelfaunan vilket gör det nödvändigt att vidta åtgärder för dess överlevnad (se bilaga 1).
Studier har gjorts med utplacering av bibatterier i naturen eller i ett slutet system (växthus) för att mäta preferens skugga-sol (Gaston et al. 2005), födosökstid i olika habitattyper (Goodell 2003, Klein 2004), födosökstid-art och födosöksavstånd-art (Gathman & Tscharntke 2002), tomma ytterceller-parasitism (Münster-Swendsen & Calabuig 2000), beskrivningar av bon i naturligt tillstånd-artificiella bon (Harris 1994), beskrivning av ny art (Paini 2004), habitatkvalitet/degradering-artförluster (Klein 2002), fluktuation av antal-tid på säsong (Horn 2003, Loyola & Martines 2006) och
orienteringbeteende runt boet (Fauria 2002). Syftena liksom metoderna är många. Genomgående beskrivs ett halvsocialt beteende där många steklar av samma eller olika art anlägger bon intill varandra.
Preferenser för håldiametrar har ofta utretts (Harris 1994, Budriene et al. 2004) och det visar sig att man kan dela in bibatteristeklarna i funktionella grupper efter vilka diametrar som används (Budryene et al. 2004). Somliga har bred diametertolerans medan andra har snäv tolerans och då främst åt
ovanliga diametrar kanske för att minska mellanartskonkurrensen om bohål. Eventuellt är preferensen kopplad till utgångshålen efter specifika skalbaggar, där snäv tolerans skulle representera ett beroende av ett litet antal skalbaggsarter (Budryene et al. 2004). Solitära steklar har stora tidsmässiga och energetiska kostnader för att leta upp lämpligt bohål (Klein et al. 2004). Eftersom det troligen är ont om bohål i naturen tvingas individerna att vara opportunistiska och acceptera de hål som finns tillgängliga.
Hur ser mönstren i boplatsval ut och vilka är orsakerna till eventuella mönster? Karakteristiskt för hålbyggande solitära steklar är beteendet att mer eller mindre intra- och interspecifikt aggregera vid boplatserna. Evolutionära reproduktionsfördelar med aggregation skulle kunna vara att, trots att sannolikheten för parasitering av populationen ökar, så blir parasittrycket för den enskilda individen mindre (Inman & Krebs 1987) och detta har visats hos skalbaggar (Morrison & Strong 1981) och hos solitära steklar (Larsson 1986). Evolutionära fördelar med att vara i centrum av en aggregation eller på en yta borde bero på situationen, exempelvis mitt på ett hygge jämfört med mitt i en homogen skog. Man kan urskilja andra strategier eventuellt kopplat till att undvika parasitering och predation hos solitära steklar, till exempel att bobygget börjar längst in i ett hål som är så litet som möjligt, att
attrapphål anläggs runt ett verkligt bohål (personlig observation), att eventuellt en mörk bakgrund väljs före ljus (personlig observation), att en tom cell anläggs ytterst, att ett bebyggt hål avslutas med propp längre in (personlig observation).
Min undersökning gjordes för att ta reda på om det fanns spatiala mönster i solitära steklars boplatsval i bibatterier. I undersökningen har alla förekommande arter sammanslagits för att urskilja generella tendenser för dessa steklar.
Material och metoder
Undersökningen utfördes 30 april – 30 augusti 2006 i landskapet Sörmland, Sverige, 20 km söder om Eskilstuna, Södermanland. Detta är en skogs- och jordbruksregion i Mälarmården. Två lokaler valdes ut, lokal 1: N 59º 15' 19.99" E 16º 29' 8.86" och lokal 2: N 59º 16' 57.94" E 16º 27' 29.22". Lokalerna är belägna 1,5 km från varandra och är före detta betesmark. Uppskattningsvis upphörde betet runt år 2000. I anslutning till lokal 1 fanns en ca 50x50 m stor ängsmark. Det fanns gott om bördig
ruderatmark på lokal 2. Delar av lokal 1 var något sank medan lokal 2 var torr och låg på en kulle. Platsen för undersökningen ligger innanför 5 mils-radien från det stora kraftvärmeverket i Eskilstuna vilket innebär att alla avverkningsytor inom området finkammas på rester av tall, gran och asp, och att även stormfällt eller skadat virke (ihåliga eller avbarkade gamla granar, björkar, aspar, och tallrakor tas om hand för att användas som (bio-)bränsle.
Till försöksuppställning konstruerades sex bibatteri-stationer av grovt virke. Stationerna skruvades ihop till homogena block med måtten 60x60x20 cm (bredd x höjd x djup) och försågs med ben ca. 5 cm höga för att komma upp en bit från marken samt ett tak med överhäng av plåt och tunn tjärpapp till skydd mot regn. Den yta i vilken bohål skulle borras hyvlades först, därefter borrades hål i ett
regelbundet mönster (se Figur 1) som upprepades i tänkta 10x10 cm rutor, allt som allt 25 sådana rutor per bibatteri. Varje hål av samma diameter lades i grupper bredvid varandra inom rutorna. Antalet hål i varje ruta var 9 st 2 mm, 6 st 3 mm, 6 st 4 mm, 6 st 5 mm, 4 st 6 mm, 4 st 7 mm, 4 st 8 mm. Således fanns totalt 975 hål i varje station. Varje hål var 14 cm djupt utom 2 och 3 mm hålen, som var 3 respektive 8 cm djupa.
Figur1. Borrhålsmönstret där varje håldiameter är placerade i grupper i tänkta 10x10 cm rutor. Bilden är ett utsnitt av en (50x50) station
Hålens stora djup var avsedda att få stekelhonorna att använda färre bohål så att studien blev mer inriktad på olika honors beteende. Alla hål försänktes. Utplaceringen skedde den 30 april på två valda lokaler. Fyra stationer på lokal 1 och två stationer på lokal 2. Då de placerades med ett avstånd på 50 - 200 m från varandra valde jag att betrakta varje station som oberoende av de andra (en stekelindivid förmodades inte ha kontakt med mer än en station). Hålöppningarna vändes mot söder.
Stationerna besöktes med ca 1 veckas mellanrum. Då mättes varje nytt bohåls position i x och y- led i mm från nedre vänstra hörnet på den aktiva ytan. Förutom position noterades också
förslutningsmaterial, håldiameter och eventuell parasitering efter förslutning. När en stekel färdigställt ett bo försluter den hålet med lera. Att en förslutning blivit parasiterad kan ses på att den är
genomstucken eller uppgrävd. Hål där förslutningar gjordes med enbart sågspån var svåra att identifiera som stekelbon och uteslöts från analyser. Likaledes uteslöts vissa bohål på grund av märkligheter såsom att förslutningarna hålen inte var kompletta. Totalt uteslöts 21 observationer av dessa anledningar.
Beskrivningar och tester av spatiala mönster i hålutnyttjandet gjordes med hjälp av ett antal speciellt framställda QuickBasicprogram. I ett första steg dokumenterades aggregationstendens genom att kalkylera avstånd till närmaste granne, det vill säga avståndet mellan ett fokalt ockuperat hål och det närmast belägna ockuperade hålet. Medelavståndet till närmaste granne inom varje station
kalkylerades, samt kalkylerades medelvärdet på dessa medelavstånd.
I ett andra steg testades aggregationstendensen genom att det aktuella programmet slumpvis distribuerade bon på befintliga hål. Slumpningarna gjordes stationsvis och inom varje station slumpades ut samma antal bon som stationen i fråga hade i verkligheten. Därefter kalkylerades, likt ovan, medelavståndet till närmaste granne inom varje station, samt medelvärdet på dessa
medelavstånd. Programmet upprepade detta 100 gånger i en montecarloprocedur och jämförde varje gång det slumpade medelavståndet inom varje station med det verkligen medelavståndet inom stationen, samt jämförde programmet varje gång medelvärdet på medelavstånd med det verkligen funna medelvärdet på medelavstånd. Härvid kunde (den verkliga) aggregationstendensen inom varje station, samt det övergripande aggregationsmönstret (alla stationer) testas statistiskt genom att se hur ofta det slumpade värdet var mer extremt eller lika extremt som det verkligt funna värdet. Som
klargörande: om endast två av 100 slumpningar resulterade i lika extremt eller mer extremt värde som det faktiskt funna var P = 0.02.
I ett tredje steg dokumenterades central tendens genom att kalkylera avståndet från ett fokalt ockuperat hål till stationens centrala punkt. Medelavståndet inom varje station kalkylerades, samt kalkylerades medelvärdet på dessa medelavstånd. I ett fjärde steg testades den centrala tendensen genom att
återigen slumpvis distribuera bon på befintliga hål. Slumpningarna gjordes stationsvis och inom varje station slumpades samma antal bon ut som stationen i fråga hade i verkligheten. Därefter kalkylerades medelavståndet till stationens centrala punkt inom varje station, samt medelvärdet på dessa
medelavstånd. Programmet upprepade detta 100 gånger i en montecarloprocedur och jämförde varje gång det slumpade medelavståndet inom varje station med det verkligen medelavståndet inom varje station, samt jämfördes varje gång medelvärdet på medelavstånd med det verkligen funna medelvärdet på medelavstånd. Härvid kunde, på samma sätt som beskrivits ovan, den (verkliga) centrala tendensen inom varje station, samt inom alla stationer testas statistiskt.
I ett femte steg beskrevs och testades andra spatiella tendenser som verkade föreligga i materialet. Metoden var återigen att inledningsvis beskriva mönstret i fråga genom att kalkylera det verkliga medelvärdet inom varje enskild station och att kalkylera medelvärdet på medelvärdena, samt att avslutningsvis statistiskt testa det funna mönstret genom 100 montecarloslumpningar liknande
proceduren beskriven ovan.
Slumpningarna gjordes i samtliga ovan beskrivna fall utan återläggning, det vill säga på ett sådant sätt varje hål bara kunde erhålla ett utslumpat bo.
Resultat
Efter bortfall av 21 oklara fall (se Material och metoder) ingick totalt 283 bebyggda bon i analyserna. Således blev endast 4.84% av de befintliga hålen bebyggda. Av bona grävdes 8 ut och hålet övertogs av en ny bebyggare och 54 hål blev parasiterade vilket ger en parasiteringsgrad på 22.91%.
Steklarna tenderade att aggregera, d v s de verkade välja att bebygga hål i närheten av redan
ockuperade hål (Tabell 1 och Figur 3). Detta mönster var signifikant i fyra av de sex stationerna, samt övergripande, d v s när medelvärdet för alla stationer testades (Tabell 1).
Tabell 1. Aggregationstendens, avstånd till närmast belägna bebyggda bohål (närmaste granne). N är
antal bebyggda bohål.
station funnet1 förväntat2 N P3
1 2.21 5.07 28 < 0.01 2 9.32 10.54 8 0.30 3 4.79 5.84 23 0.11 4 1.47 2.26 120 < 0.01 5 2.00 3.51 56 < 0.01 6 2.11 3.82 48 < 0.01 1-64 3.65 5.14 283 < 0.01
1 Medelvärde (cm). 2 Medelvärde (cm) beräknat genom Monte Carlo simuleringar (se Metoder för fler
detaljer). 3 Beräknat genom Monte Carlo simuleringar (se Metoder för fler detaljer). 4 Medelvärde alla stationer.
Vad gäller central tendens, d v s benägenhet att bebygga hål i centrala delar av varje station, var mönstret mindre klart. Det fanns en signifikant sådan tendens i en station (Station 4, se Tabell 2), men detta mönster förelåg inte i någon av de andra stationerna och inte heller som övergripande mönster (Tabell 2).
Tabell 2. Central tendens, avstånd till stationens mittpunkt. N är antal bebyggda bohål.
station funnet1 förväntat2 N P3
1 19.62 19.29 28 0.57 2 17.68 19.01 8 0.30 3 17.08 19.35 23 0.08 4 18.12 19.34 120 0.01 5 20.03 19.30 56 0.87 6 20.69 19.37 48 0.92 1-64 18.87 19.08 283 0.37
1 Medelvärde (cm). 2 Medelvärde (cm) beräknat genom Monte Carlo simuleringar (se Metoder för
fler detaljer). 3 Beräknat genom Monte Carlo simuleringar (se Metoder för fler detaljer).
4 Medelvärde alla stationer.
Märkligt nog verkade det finnas en tendens att bebygga hål till vänster om mittlinjen på
stationerna. Detta mönster var signifikant i två av stationerna, samt var det övergripande mönstret signifikant (Tabell 3, se även Figur 4).
Tabell 3. Speciella spatiala mönster – vänstertendens, andel bebyggda bon vänster om mittlinjen på
varje station. N är antal bebyggda bohål.
station funnet1 förväntat2 N P3
1 78.57 50.39 28 < 0.01 2 75.00 50.75 8 0.11 3 52.17 50.16 23 0.49 4 51.67 51.08 120 0.44 5 69.64 51.49 56 < 0.01 6 52.08 50.99 48 0.48 1-64 63.19 50.29 283 < 0.01
1 Medelvärde (%). 2 Medelvärde (%) beräknat genom Monte Carlo simuleringar (se Metoder för fler
detaljer). 3 Beräknat genom Monte Carlo simuleringar (se Metoder för fler detaljer). 4 Medelvärde alla stationer.
Andra märkliga spatiella boplatsvalsmönster var att det förelåg en signifikant tendens att hål ovanför mittlinjen bebyggdes på två stationer (Station 5 och 6, se Tabell 4), samt att motsatsen, d v s att hål nedanför mittlinjen bebyggdes på en station (Station 4, se Tabell 5). Det övergripande mönstret var att hål ovanför mittlinjen bebyggdes, men mönstret var inte signifikant (Tabell 4 och Tabell 5).
Tabell 4. Speciella spatiala mönster – vertikal tendens, andel bebyggda bon ovan mittlinjen på varje
station. N är antal bebyggda bohål.
station funnet1 förväntat2 N P3
1 60.71 50.39 28 0.17 2 75.00 54.25 8 0.20 3 39.13 50.46 23 0.85 4 35.00 52.17 120 1.00 5 62.50 50.41 56 0.02 6 66.67 51.09 48 0.01 1-64 56.50 51.63 283 0.10
1 Medelvärde (%). 2 Medelvärde (%) beräknat genom Monte Carlo simuleringar (se Metoder för fler
detaljer). 3 Beräknat genom Monte Carlo simuleringar (se Metoder för fler detaljer). 4 Medelvärde alla stationer.
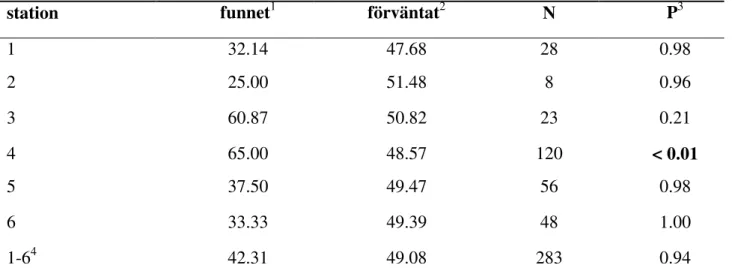
Tabell 5. Speciella spatiala mönster – omvänd vertikal tendens, andel bebyggda bon under mittlinjen
på varje station. N är antal bebyggda bohål.
station funnet1 förväntat2 N P3
1 32.14 47.68 28 0.98 2 25.00 51.48 8 0.96 3 60.87 50.82 23 0.21 4 65.00 48.57 120 < 0.01 5 37.50 49.47 56 0.98 6 33.33 49.39 48 1.00 1-64 42.31 49.08 283 0.94
1 Medelvärde (%). 2 Medelvärde (%) beräknat genom Monte Carlo simuleringar (se Metoder för fler
detaljer). 3 Beräknat genom Monte Carlo simuleringar (se Metoder för fler detaljer). 4 Medelvärde alla stationer.
0 10 20 30 40 50 Position i x-led (cm) 0 10 20 30 40 50 P o s iti o n i y -l e d ( c m )
Figur 3. Generellt ockupationsmönster, alla stationer (1-6) överlagrade. Cirklar representerar bebodda bohål 0 10 20 30 40 50 Position i x-led (cm) 0 10 20 30 40 50 P o s iti o n i y -l e d ( c m )
Figur 4. Ockupationsmönster i station 5. Cirklar representerar bebodda bohål. Tendensen att steklarna tenderade att bygga bo i aggregationer är tydlig
Diskussion
Studien visar att steklarna hade en klar tendens att aggregera, det vill säga att välja bohål nära andra ockuperade bohål (dock måste det tilläggas att det kan ha förekommit att individer bebyggt två eller flera bohål i närheten av sitt första bohål trots att hålen var djupa, efter det att individen under
bobygget observerat omgivande nära lämpliga bohål). Det ligger nära till hands att tro att anledningen till detta kan vara att de därigenom kan undvika parasitering och predation (Hamilton 1971).
Aggregationstendenser både under fortplantning och vid andra tidpunkter har påvisats hos många arter inom olika taxa t.ex fiskar, fåglar och däggdjur (Robinson 1985, Mooring & Hart 1992). Om
aggregation skedde för att steklarna försökte unvika parasitering eller predation, kunde man också ha förväntat en central tendens. I en selfish herd (Hamilton 1971) rör sig individerna mot centrum av en grupp (Viscido & Wethey 2002) och tätheten mellan individer och den centrala positionen resulterar i ökad fitness (Olmos 2003). Vissa parasiter och rovdjur går inte att skydda sig mot med hjälp av aggregation, som när t.ex parasitflugor attackerar en klövdjursflock. Flugorna attraheras av den stora flocken och lägger så många ägg som möjligt på flockens individer (Mooring & Hart 1992). Ett exempel där beläggen för den eventuella förekomsten av selfish herd går isär, är gruppbeteendet hos sandkrabban Uca pugilator. Där Viscido & Wethey (2002) funnit stöd för teorin, har McLain et al. ( 2005) funnit motsatsen. Bland kolonihäckande fåglar finns rumsliga häckningsmönster i boplatsvalen (Robinson 1985). Koloniplatsvalet kan bestämmas efter omständigheter som t.ex otillgänglighet (öar). Robinson (1985) fann för Cacicus cela, som tillhör familjen vävarstarar, att även gamla övergivna bon försvårade för predatorer på samma sätt som om kolonin var större än den var, både i
avskräckningssyfte och för möjligheten att gömma boet i mängden. Vid förstörd häckning bytte fåglarna koloni, vilket resulterade i färre men större kolonier på de lämpligaste platserna. Bon i kolonier av en ibis-art (Eudocimus ruber) som Olmos (2003) undersökte hamnade under andra
häckningen närmare varandra och hade ökad häckningframgång, som var kopplad till minskat avstånd till närmaste granne. För att återkoppla till steklarnas aggregation, fann Larsson (1986) hos en
markstekelart (Bembix rostrata) ett mönster, där botätheten var positivt korrelerad med antalet
parasiter, medan sannolikheten för varje stekelindivid att bli parasiterad minskade. För konsumenter är så gott som alltid resurserna i omgivningen fläckvis distribuerade (Begon et al. 1990). Det kan
förmodas att detta också gäller för reproduktionsplatser, i det här fallet en fläckvis ansamlad resurs med bokammare (att jämföra med lämplig sandmark för marksteklar). Steklarna är benägna att stanna kvar på sin kläckningsplats (nathal philopatry) (Polidori et al. 2006). I enlighet med Klein et al. (2004) borde kostnaderna för boplatssök minska och därmed skulle sannolikheten att hitta en boplats för individen och avkomman och dess genetiska fitness öka. I den genomförda studien var ingen stekel kläckt i stationerna, dvs. alla steklar som förekom var av första generationens kolonisering (founder) (med undantag för andragenerations-kolonisering under säsongen om det eventuellt förekom). Usurpationsbeteenden, dvs. beteenden som leder till att en individ övertar ett upptaget bohål, och konkurrensen om bohål leder till strider mellan honor ibland med dödlig utgång (Batra 1978). Man har studerat släktskap inom kolonier för att kunna bedöma vilken typ av socialitet som råder (Polidori et al. 2006). Socialitet antas här inte ha avgörande betydelse om man ska bedöma varför aggregation sker, det blir dock relevant när det är frågan om eusociala steklar inom exempelvis Halictidae (Strohm & Bordon-Hauser 2003, Wyman & Richards 2003) och Colletidae (Spessa et al. 2000) och då kan både socialitet och storlek på aggregationer variera inom arter beroende på lokala ännu obestämda ekologiska faktorer (Wyman & Richards 2003). Det fanns inte några observationer av att några eusociala steklar förekom i studien. I den här studien aggregerade steklarna trots ett överflöd av hålkammare. 4,84% av hålen användes i första generationens kolonisation. Begon et al. (1990)
förklarar begreppet födosöksaggregation, och förutom selfish herd-fördelar (minskad parasitering och predation), kan andra fördelar vara tänkbara som främjar ansamling av individer, fler partner att välja mellan vid reproduktion, förbättrad upptäckt av födoresurs, bättre födointag (till exempel när en barkborre-individ koloniserar ett träd så utsöndras aggregationsferomoner och tillsammans försvagar
individerna trädet. I studien har hittats boplatsaggregation. Solitära steklar orienterar sista biten till bohålet med hjälp av luktorgan i antennerna (Fauria et al. 2004). Det kan tänkas att detta också används för att stimulera/attrahera honor att bygga intill ockuperade bohål, även honor av andra arter skulle kunna bli attraherade. Batra (1978) rapporterar att markbiet Nomia melanderi verkar lockas till bobyggnadsbeteende endast där det finns redan anlagda bon även om det fanns gott om bra boplatser i området. När honor fångades i nät och flyttades till en annan plats samlades en svärm andra honor runt nätet. En sådan fläckvis koloni kan fortleva i många år. Både selfish herd och den fläckvisa
distributionen av reproduktionsplatser kan båda bidra till bibatteristeklars aggergationsbenägenhet. Tanken med centrum-hypotesen var att steklarna skulle bygga i mitten på bibatterierna, i enlighet med Viscido & Wethey (2002), för att därmed undvika en eventuellt existerande kant-effekt, där parasit- och rovsteklar skulle hålla till. Eller tvärt om en perifer tendens, beroende på var i
predator-byteskapplöpningen som evolutionen befinner sig för tillfället. Liksom hos fågelkolonier är hela ytan exponerad för attacker, och det kan tänkas fördelaktigt att välja en position, som parasiterna inte i första hand väljer. Parasiterna å andra sidan följer efter men kommer på efterkälken, om denna kapplöpning skulle existerat, men sinnesorgan som syn och lukt är så viktiga hos predatorer att de kopplar förbi instinkter och går direkt på det nya bytet/bohålet. Trädkolonifåglar är mer utsatta för predation i en miljö där kolonin har en kantzon där bon får utstå attacker på grund av kanteffekten, medan en koloni i homogen skog inte har denna kantpredation (Robinson 1985). I båda situationerna är kolonierna exponerade för rovfåglar ovanifrån, men ju större kolonin är desto fler individer väntas mobba anfallaren och avstyra eller försvåra angreppet (Olmos 2003). En annan aspekt utesluter inte att en centrumtendens för de enskilda aggregationerna av bohål på bibatteriet förekom i en mindre skala än den som analyserades utifrån hela stationens boyta (skalan på vilken steklarna och deras fiender lever på, är förmodligen mindre än stationens mått 50x50 cm). Där skulle de bokammare som ligger i mitten av varje koloni kunna få en lägre parasiteringsgrad. Men vilka är kriterierna för detta? Förutom usurpation kan steklarna inte välja i mitten om hålen är upptagna, utan bara i tomma hål intill
ockuperade hål. Det finns alltså två alternativ, antingen att vara founder/usurpator eller lägga sig intill tidigare steklars hål. Selektion skulle då grunda sig på faktorn tid. Tidig ägglägning- högre genetisk fitness. Och för varje bohål som honor anlägger skulle då öka reproduktionsframgången inte i direkt proportion till antalet lagda ägg utan mera exponentiellt. Parasitsteklar (ej clepto-) arbetar långt efter att värdsäsongen avslutats (personlig observation). Var bohålets placering i aggregationen av någon betydelse? Eller är aggregationsfördelarna oberoende av placering? Megachile-arter, som arbetar senare på säsongen än Osmia-arter, möter förmodligen färre lediga boplatser, men att tidsaspekten på boplatssituationen skulle ge dem lägre reproduktiv framgång är inte troligt (hellre i så fall sämre väderleksförhållanden såsom väta och lägre temperatur på sensommaren). En observation (Bild 2) som kan bekräfta en här formulerad misstanke om att (koloni)centrumteorin inte stämmer, är att hål
belägna ytterst används trots att det innebär att de alltid kommer att vara ytterst i en aggregerad koloni. Tyvärr var en analys i mindre skala inte möjlig att genomföra i studien.
Figur 2. Aggregation av minst tre arter. 5 bohål har valts med perifer position. Bilden har retusherats för att synliggöra de 4 sammanhängande aggregationerna. Bibatteriet på bilden ingår ej i analysdata
Vänstertendensen dvs. att steklarna verkade välja boplatser i vänstra delen av stationen är ett intressant men svårtolkat resultat. Någon förklaring till varför är svår att ge. Litteraturen ger exempel på att det observerats tidigare, men ger inte några ledtrådar till förklaring. Alla stationer var orienterade på samma sätt vättande mot söder. Det kan tänkas att mönster i förhärskande vindriktning, riktning till solens uppgång, solexponering och mikroklimat och liknande faktorer kan ha haft betydelse för boplatsvalet.
Referenser
Banaszak J., 1992, Strategy for conservation of wild bees in an agricultural landscape, Agriculture,
Ecosystems and Environment, 40:179-192.
Batra S. W. T., 1978 Aggregation, Territoriality, mating and nest aggregation of some solitary bees (Hymenoptera: Halictidae, Megachilidae, Colletidae, Anthophoridae), Journal of the Kansas Entomological Society, 51:547-559.
Begon M., Harper J. L. & Townsend C. R., 1990, Ecology: individuals, population and comunities,
second edition, Blackwell Scientific Publication, 298-334.
Budriene A., Budrys E. & Nevronytė Ž., 2004, Solitary Hymenoptera Aculeata inhabiting trap-nests in Lithuania: nesting choice and niche overlap, Latvijas Entomologs, 41:19-31.
Cane J. H. & Tepedino V. J., 2001, Causes and extent of declines among native north american invertebrate pollinators: detection evidence, and consequences, Conservation Ecology, 5, (online).
Donovan B. J., 1980, Interactions between native and introduced bees in New Zealand, New Zealand
Journal of Ecology, 3:104-116.
Ehnström B. & Axelsson R., 2002, Insektsgnag i bark och ved, Artdatabanken SLU, Uppsala. Fauria K., Campan R. & Grimal A., 2004, Visual marks learned by the solitary bee Megachile
rotundata for localizing its nest, Animal Behaviour, 67:523-530.
Gaston K. J., Smith R., Thompson K. & Warren P. H., 2005, Urban domestic gardens (II): experimental tests of methods for increasing biodiversity, Biodiversity and Conservation, 14:395-413.
Gathmann A. & Tscharntke T., 2002, Foraging ranges of solitary bees, Journal of Animal Ecology,
Gess F. W. & Gess S. K., 1999, The use by wasps, bees and spiders of shells of Trigonephrus Pilsb. (Molusca: Gasteropoda: Dorcasiidae) in desertic winter-rainfall areas in southern Africa, Journal of Arid Environments, 43:143-153.
Goodell K., 2003, Population ecology: Food availability affects Osmia pumila (Hymenoptera:
Megachilidae) foraging, reproduction, and brood parasitism, Oecologia, Springer- Verlag.
Granvik H., 1928, Våra fåglars nytta och skada, Naturens liv i ord och bild, andra delen, Bokförlaget
Natur och kultur.
Hamilton W. D., 1971, Geometry of the selfish herd, J. theor. Biology, 31:295-311.
Inman A. J. & Krebs J., 1987, Predation and group living. Trends in Ecology and Evolution,
2:31-32,
Klein A.-M., Steffan-Dewenter I. & Tscharntke T., 2004, Foraging trip duration and density of megachilid bees, eumenid wasps and pompilid wasps in tropical agroforestry systems, Journal of Animal Ecology, 73:517-525.
Kreuss A. & Tscharntke T., 2002, Grazing intensity and the diversity of grasshoppers, butterflies, and trap-nesting bees and wasps, Conservation Biology, 16:1570-1580.
Kullenberg B., 1949, Insekter och blommor, Svenska djur: Insekterna, Bohuslänningens AB,
Uddevalla, 65-83.
Larsson F. K., 1986, Increased nest density of the digger wasp Bembix rostrata as a responsa to parasites and predators (Hymenoptera: Sphecidae), Entomol. Gener., 12:071-075.
Larsson M., 2006, To bee or not to be: critical floral resources of wild-bees, Digital Comprehensive
Summaries of Uppsala Dissertations from the Faculty of Science and Tecnology 210, Uppsala.
Linkowski W. I., Cederberg B. & Nilsson L. A., 2004, Vildbin och fragmentering:
kunskapssammanställning om situationen för de viktigaste pollinatörerna i det svenska jordbrukslandskapet, Svenska Vildbiprojektet vid Artdatabanken, SLU och Avdelningen för
växtekologi, Uppsala universitet, Uppsala.
Loyola R. D. & Martines R. P., 2006, Trap-nest occupation by solitary wasps and bees (Hymenoptera: Aculeata) in forest urban remanent, Neotropical Entomology, 35:41-48.
McLain D. K., Pratt A. E. & Kirschstein K., 2005, Predator-driven fragmentation of fiddler crab droves into selfish miniherds of biased composition, Journal of Experimental Marine Biology and Ecology, 315:1-15.
Mooring M. S. & Hart B. L., 1992, Animal grouping for protection from parasites: selfish herding and encounter-dilution effects, Behaviour, 123:173-192
Morrison G. & Strong D. R., 1981, Spatial variations in egg density and the intensity of parasitism in a neotropical chrysomelid (Cephaloleia consanguinea), Ecological Entomology, 6:55-61
Münster-Swendsen M. & Calabuig I., 2000, Interaction between the solitary bee Chelostoma
florisomne and its nest parasite Sapyga clavicornis- empty cells reduce the impact of parasites, Ecological Entomology, 25:63-70.
Nilsson I., 2004, Betesdriftens betydelse för mångfalden av kärlväxter, insekter och spindlar,
opublicerat, Mälardalens högskola, Eskilstuna.
Olmos F., 2003, Nest location, clutch size and nest success in the Scarlet Ibis Eudocimus ruber, Ibis,
145:E12-E18, (online).
Paini D. R. & Roberts J. D., 2005, Commercial honey bees (Apis mellifera) reduce the fecundity of an australian native bee (Hylaeus alcyoneus), Biological Conservation, 123:103-112.
Paini D. R., 2004, Nesting biology of an australian resin bee (Megachile sp.; Hymenoptera:
Megachilidae): a study using trap nests, Australian journal of entomology, 43:10-15.
Pettersson B., 2003, Vilda bin och solitära getingar- trädgårdens bundsförvanter, Fakta
Trädgård-fritid, Sveriges lantbruksuniversitet, Uppsala.
Polidori C., Casiraghi M., Di Lorenzo M., Valarani B. & Andrietti F., 2006, Philopatry, nest choice, and aggregation temporal-spatial change in digger wasp Cerceris arenaria
Quaranta M., Ambroselli S., Barro P., Bella S., Carini A., Celli G., Cogoi p., Comba L., Comboli R., Felicioli A., Floris I., Intoppa F., Longo S., Maini S., Manino A., Mazzeo G., Medrzycki P., Nardi E., Niccolini L., Palmieri N., Patetta A., Piatti S., Piazza M. G., Pinzauti M., Porporato M., Porrini C., Ricciardelli D´Albore G., Romagnoli F., Ruiu L., Satta A., Zandigiacomo P., 2004, Wild bees in agroecosystems and semi-natural landscapes. 1997-2000 collection period in Italy, Bulletin of Insectology, 57:1:11-61.
Robinson S., 1985, Coloniality in the Yellow-rumped Cacique as adefence against nest predators, The
Auk, 102:506-519.
Samuelsson J. & Ingelög T., 1996, Den levande döda veden- bevarande och nyskapande i naturen,
ArtDatabanken, SLU, Uppsala, 26-28.
Sandhall Å. & Lindroth C. H., 1998, Skalbaggar: utseende utveckling levnadssätt beteende, andra
upplagan, Stenströms bokförlag/Interpublishing, Stockholm.
Spessa A., Schwarz M. P. & Adams M., 2000, Sociality in Amphylaeus morosus (Hymenoptera:
Colletidae: Hylaeinae) Annals of the Entomological Society of America, 93:3:684-692.
Strohm E. & Bordon-Hauser A., 2003, Advantages and disadvantages of large colony size in a halictid bee: the queen´s perspective, Behavioral Ecology, 14:546-553.
Tscharntke T., Gathmann A. & Steffan-Dewenter I., 1998, Bioindication using trap-nesting bees and wasps and their natural enemies: community structure and interactions, Journal of Applied
Ecology, 35:708-719.
Viscido S. V. & Wethey D. S., 2002, Quantitative analyses of diddler crab flock movement: evidence for ‘selfish herd’ behaviour, Animal behaviour, 63:735-741.
Williams I. H., 2002, Insect pollination and crop production: a european perspective, sid. 59-65 i:
Kevan P & Imperatriz Fonseca VL (eds) - Pollinating Bees - The Conservation Link Between Agriculture and Nature – Ministry of Environment/Brasília.
Wyman L. M. & Richards M. H., 2003, Colony social organisation of Lasioglossum melachurum Kirby (Hymenoptera, Halictidae) in southern Greece, Insect Sociaux, 50:201-211.
Bilaga 1. Hot mot stekelfaunan
I östra Europa har ett extremt högt antal biarter (i ungefärlig storleksordning 500-800) kunnat klara sig på jordbruksmarkerna beroende på att man där endast använder viss andel av de öppna fälten till åkrar (Banaszak 1992). Resten används till bete, slåtter eller annat mindre intensivt jordbruk och många fält ligger i ständig träda. Steklarna som studerats i mitt projekt är på gränsen till fortsatt existens i vårt land och i övriga Europa. Mängden solitära steklar begränsas i första hand av tillgången på boplatser (Williams 2002). Tillgången på boplatserna är i sin tur beroende av mängden död och döende ved på både skogsbruks- och jordbruksmark. Lämplig död och döende ved, måste för att kunna användas av steklar som boplatser primärt ha skapats av skalbaggar som gör gångsystem. Barkborrearter jämte brand, storm, torka eller regn som ger svamp- och insektsangrepp kan bidra med naturlig succession, där sekundära vedlevande arter gynnas. En stor mängd skogslevande skalbaggar har fått sin utbredning starkt beskuren. Vissa urskogsarter har inte hittats i Sverige på mer än hundra år. Reglering av vatten i inlandet och skogsbrukets metodiska avlägsnande av gamla murkna träd i skogen tros ligga bakom (Sandhall & Lindroth 1998). Skalbaggars gångsystem och utgångshål har en direktkoppling till stekelpopulationerna, även om dessa måste vara i gränsmarken mellan skog och öppen yta för att kunna återanvändas som bokammare. Man skulle kunna dra en parallell till en snarlik
beroendesituation i naturen nämligen hålhäckande fåglars (och gnagares) användning av bokammare uthackade av hackspettar och ihåligheter skapade av svampar, hästmyror m.fl. (Samuelsson & Ingelög 1996). Resurserna måste alltid nyskapas i överflöd och variation. När skogarna rensas på det döda eller framtida döda virket för transport till fjärvärmeverken under namnet förnyelsebar energi så är det förstås inte bara dessa insektspopulationer som blir lidande, det medför en direkt påverkan även på svampar, fåglar, groddjur, reptiler samt lägre och högre däggdjur.
En andra begränsande faktor är tillgången på blomrika ängar, ruderatmark och växter på åkrarna vilket ger nektar och pollen och värdväxter för bytesinsekter. Idag finns en hel del redan utdöda arter i Sverige (Ehnström & Axelsson 2002). Öppna fält skapades först av de vilda betesdjuren, sedan av jordbrukets slåtter och bete, och nu utgör igenväxning av betes- och slåttermark samt omvandlingen till skogsbruk och storskaligt jordbruk ett stort hot mot insekter generellt (Nilsson 2004). Dock skall sägas att vad gäller betet så har det visat sig att ett mindre intensivt bete ökar biodiversiteten i ett område och det kan vara avgörande att tänka på detta i restaureringsarbeten (Kreuss & Tscharntke 2002). Där betesdrift och ängshävd tidigare knutit samman större områden kan nutida splittrade rester av populationer avskiljda från varandra påtvingas inavel.
Till dessa faktorer tillkommer att stigande ålder på habitaten ökar biodiversiteten. Alla dessa faktorer påverkas negativt när gammelskog på väg mot urskog byts ut mot
produktionsskog, åkrar breder ut sig och urban miljö växer, så att på områden där tidigare smålantbruk legat biarter formligen grävs ur slänter och blottad mark till förmån för stadens utbyggnad.
Populationer som valt att bosätta sig i ett dött träd på bondens mark var tidigare fredade men hamnar nuförtiden i fjärrvärmeverket eller blir pappersmassa.
I vissa områden, som till exempel i Australien, på Nya Zeeland och i Sydamerika har man medvetet introducerat främmande arter solitärbin och getingar i stället för att gynna redan befintliga
populationer (Donovan 1980). Det har också blivit populärt att odla kulturer av inhemska arter (Bombus spp. och Osmia spp.) i Europa och Nordamerika. Vad kommer det att få för följder för de naturliga bestånden? Tambiet har visat sig konkurrera med vilda bin om nektar och pollen och det måste man väga in i restaureringsbilden (Linkowski et al. 2004, Paini & Roberts 2005).