1
Department of Physics, Chemistry and Biology
Master Thesis
How does predation from fish influence the benthic
invertebrates’ species composition in the Phragmites
australis and Chara vegetation of Lake Takern?
Ernest Aigbavbiere
LITH-IFM-A-EX--10/2310—SE
Supervisor: Anders Hargeby, Linköpings universitet
Examiner: Karin Tonderski, Linköpings universitet
Department of Physics, Chemistry and Biology
Linköpings universitet
2
IFM, Linkoping University, Linkoping , Sweden.
Rapporttyp Report category Licentiatavhandling x Examensarbete C-uppsats D-uppsats Övrig rapport _______________ Språk Language Svenska/Swedish x Engelska/English ________________ ISBN ______________________________________________ ____ ISRN ______________________________________________ ____
Serietitel och serienummer ISSN
Title of series, numbering
LITH-IFM-A-Ex—10/2310-SE
Datum
Date 2010-05-28
URL för elektronisk version
Sammanfattning
Abstract
Predation is one of the important selective factors that regulate the species composition of benthic invertebrate communities. The study objective was to investigate the invertebrate distribution in two contrasting habitats in Lake Takern, southern Sweden, submerged Chara vegetation and emergent Phragmites australis vegetation, and to investigate the influence of predation from fish on certain invertebrates. Laboratory studies were used to estimate handling time and the intake rate (mg/s) by the fish based on the optimal foraging model. In the field, fish and invertebrates were collected with gill nets and hand nets respectively and the fish gut content was analyzed. In total, sixteen invertebrates’ taxa were collected from the two habitats. The proportion of the invertebrate’s overlaps from each of the habitat was calculated by Renkonen index and with Sorensen similarity index. Both indices showed a similarity larger than 65%, indicating that there was no significant difference in the invertebrates’ distribution in the reed and the Chara habitat. The fish caught with the gill nets were: roach (Rutilus rutilus), perch (Perca fluviatilis),tench (Tinca tinca), and rudd (Scardinius
erythrophthalmus). The caught perch had eaten: Asellus aquaticus, Gammarus lacustris, Corixidae, and the larvae of Chironomidae and Zygoptera.A comparison was made on the invertebrates found in the field and the ones observed from
the gut of the perch and the findings were that the invertebrates that had more occurrence in the gut were less in proportion in the benthic samples. In the laboratory experiment perch ( Perca fluviatilis)) was used as the predator fish and the prey organisms were Asellus aquaticus,Gammarus pulex, and Corixidae of three size categories. The results showed that perch handling time for A.aquaticus of the different size categories, was not significantly different (p>0.05); and the same results were valid for Corixidae and G. pulex. However, the intake rate of perch across the prey and their size categories were significantly different. The handling time was not significantly different which means that the predator fish will gain more in terms of intake rate as it feeds on larger size prey items, thus harmonizing with the optimum foraging theory.
Nyckelord
Keyword: Handling time, intake rate, predation, macro invertebrates.
Titel
Title
How does predation from fish influence the benthic invertebrates’ species composition in the
Phragmites australis and Chara vegetation of Lake Takern?
Författare
Author Ernest Aigbavbiere
Avdelning, Institution
3
Content
1 Abstract... 4
2 Introduction..…..……….……… ... 6
3 Materials and Methods.. .…..….………... 7
2.1 Description of the study site... 7
2.2 Sampling of fish and invertebrates... 8
2.3 Experimental design... 9
2.4 Data and data analysis... 9
4 Results.….…….…..………... 10
3.1 Fish sample... 10
3.2 The benthic invertebrates samples... 11
3.3 Fish gut content ... 12
3.4 Fish predation experiment... 12
5 Discussion.….….….………... 17
4.1 Sampling type ... 17
4.2 Fish predation on invertebrates... 18
4.3 The results and the research questions... 18
4.4 Summary and conclusion... 20
6 Acknowledgement... 20
7 References……….. ………….... 21
4
Abstract
Predation is one of the important selective factors that regulate the species composition of benthic invertebrate communities. The study objective was to investigate the invertebrate distribution in two contrasting habitats in Lake Takern, southern Sweden, submerged Chara vegetation and emergent Phragmites australis vegetation, and to investigate the influence of predation from fish on certain invertebrates. Laboratory studies were used to estimate handling time and the intake rate (mg/s) by the fish based on the optimal foraging model. In the field, fish and invertebrates were collected with gill nets and hand nets respectively and the fish gut content was analyzed. In total, sixteen invertebrates’ taxa were collected from the two habitats. The proportion of the invertebrate’s overlaps from each of the habitat was calculated by Renkonen index and with Sorensen similarity index. Both indices showed a similarity larger than 65%, indicating that there was no significant difference in the
invertebrates’ distribution in the P. australis and the Chara habitat. The fish caught with the gill nets were: roach (Rutilus rutilus), perch (Perca fluviatilis), tench (Tinca tinca), and rudd (Scardinius erythrophthalmus). The caught perch had eaten: Asellus aquaticus, Gammarus
lacustris, Corixidae, and the larvae of Chironomidae and Zygoptera.A comparison was made
on the invertebrates found in the field and the ones observed from the gut of the perch, and the findings were that the invertebrates that had more occurrence in the gut were less in
proportion in the benthic samples. In the laboratory experiment perch ( Perca fluviatilis) was used as the predator fish and the prey organisms were Asellus aquaticus,Gammarus pulex, and
Corixidae of three size categories. The results showed that perch handling time for
A.aquaticus of the different size categories, was not significantly different (p>0.05); and the
same results were valid for Corixidae and G. pulex. However, the intake rate of perch across the prey and their size categories were significantly different. The handling time was not significantly different which means that the predator fish will gain more in terms of intake rate as it feeds on larger size prey items, thus harmonizing with the optimum foraging theory. Keywords: Handling time, intake rate, predation, macro invertebrates.
5
1 INTRODUCTION
Predation is attributed as a major selective force structuring the benthic invertebrate communities (Wellborn et al.1996). It has also been shown that habitat differences and its gradients may influence such predation and the community structure. The freshwater community of shallow lakes include species of fish, numerous invertebrates, amphibians, some reptiles and birds. Some invertebrates are known to be associated with some types of submerged vegetation and have evolved some phenotypic expressions in terms of their sizes and pigmentation. The predatory activities of the fish on invertebrates is perceived to be limited in areas rich in aquatic plants (Gilinsky 1984; Diehl 1992; Beckett et al., 1992); and the reverse in open areas (Tolonen et al., 2003). Fish is considered a top predator in most permanent aquatic systems. They exhibit high preference for large invertebrates, prey that swims actively; while predatory invertebrates may have preference for less active, small size preys (Wellborn et al., 1996). To this ends, environments with high fish predation will in turn promote selection (Nilsson 1981; Crowder and Cooper 1982; Bechara et al. 1993; Blumenshine et al. 2000).
Many studies have been conducted in looking at the combined influences of fish and invertebrates predators on the community structure of benthic macro invertebrates. Most of such investigations are focused on interactions in flowing water systems. Shallow lakes or ponds provide suitable spots to investigate more closely how the two predation pressures can affect the community structure of the aquatic macro invertebrates. To understand the phenomenon of predation some theories have been advanced by researchers to describe the processes that modulate predator and prey relations. One of such theories is the optimal foraging concept (Emlen 1966; MacArthur and Pianka, 1966; Charnov 1976; Mittelbach 1981). It’s proposed that, in selecting prey size, predators weigh the cost of handling a prey, against the benefits derivable in terms of energy gain from the consumption of the prey; a predator will select prey size that will generate maximal energy uptake per unit of handling time (Smallegange et al. 2008). The adequacy of this theory was demonstrated in many studies involving molluscivores predators (Elner and Hughes 1978; Hughes and Seed, 1995; Mistri 2004). However, these foragers on mollusc also express a flexible approach to prey selection, by feeding on sizes that are less energy giving in order to reduce the cost or risk incurred during the handling, or as a response to hunger, presence/absence of a competitor; this described another theory which is called contingency theory (Hughes 1988; Visser 1995; Johnstone and Norris, 2000; Rutten et al. 2006). Optimal foraging theory has become an important tool in studying feeding ecology because of its quantitative appeal in the prediction of predator – prey relationship (Persson and Greenberg 1990; Jackson and Rundle 2008). Essentially, the optimal foraging model is to maximize E/t; where E is the predator net energy gain from the consumption of the prey, and t, the time taken for handling and searching its prey. The cost also includes the risk incurred in the handling of the prey; so there is the question of the prey profitability, when different preys are available to the predator at the same time. Thus Jackson et al. (2004) and Jackson and Rundle (2008) adapted the net energy optimal foraging model to describe the diet shifts of goby (Pomatoschistus microps), when a variety of prey was available.
6
………. i
Where E (J), t (s), λi rate of predation on prey i (items s−1), A is assimilable part of energy, ei is
energetic content of prey i (J), Ch is handling cost (J s
−1
), Hi is handling time for preyi (s) and
Cs, the cost of searching (J s−1). A, assumed constant at 0.7, according to Elliott (1976). The
results of the study demonstrated among others that an increase in prey size for goby correlated with an increase in the handling time and also that the predator size related directly to the size of the prey they consumed.
1.1 RESEARCH OBJECTIVES
The objectives of the research were: (i) to identify the fish species and the macro-invertebrates present in the studied area, (ii) to find out the predator- prey relationship between the fish and the invertebrates, (iii) to find out the differences in frequency of invertebrates occurrence in the environment as compared to the feeding; and (iv) to relate the findings to the optimal foraging model, more specifically relating it to the prey intake rate. 1.2 HYPOTHESIS:
Ho There is no significant differences in macro invertebrates’ species compositions and their frequencies in the Phragmites australis and Chara vegetation in Lake Takern.
Ho There is no relationship between the proportion of prey found in the fish gut and the fish handling time for the prey organisms.
7
2 MATERIALS AND METHODS
2.1 DESCRIPTION OF THE STUDY SITE
Lake Tåkern is located in the south of Sweden in Ostergotlands provincial district, 70km west of Linkoping. The lake attracts numerous birds and is famous as a bird lake. It spans across an area of 12km long and 8km wide, with depth averaging 0.8metres. The lake and its contiguous area are a nature reserve since 1975 and it is part of Ramsar Convention list of protected nature. The site remains an enviable resource for nature conservation, tourism and research. The nature reserve is managed by both private ownership and the state and about 5400 hectares is the size of the reserve area. 270 bird species have been found feeding and nesting in the area by bird watchers and other visitors. Agriculture, livestock graze land and forest tree productions are the common land uses in the basin.
The lake habitat is structured by the type of vegetation present. At the P.australis, there are aquatic grasses dominating the landscape and with a huge deposit of organic matter. Further, there are clear water areas with sand and mud bottom, also areas with submerged vegetation distinctly, stonewort (Chara spp) and water milfoil (Myriophyllum spicatum). The Chara dominated area has also clear water, as the Chara is seen some few centimeters under the water, while the Myriophyllum are seen up to the water surface.
Figure 1. Map of Lake Tåkern. The white dots represent the areas where sampling took place: Saby and Glanas geographical positions of the lake. The P australis areas are represented by two white dots while Chara by only one dot (Google earth 2009).
Different approaches had been used by researchers in a bid to understand the complex nature of interaction in an ecological community. Some seems to narrow investigation to a particular population, while others may prefer a mix of populations. In shallow lakes, the use of cages or similar barriers to partition experimental block or units is well accepted (Bronmark 1994). However, such block or partitioning may occur in nature as exemplified by the vegetative landscape of the studied area. Also, common is the use of core sampling device to get the invertebrate community in the mud bottom in the studied area. There is also the possibility of making experimental set up with plastic bowl or aquaria and having different substrates bottom and introducing the predator and the prey.
8 2.2 SAMPLING OF FISH AND INVERTEBRATE
The approach I adopted in the study was a combination of both field sampling and a laboratory investigation. At Lake Takern, the benthic invertebrates were sampled with a standardized 1 meter diameter wooden handle sweep net of 35cm diameter. Two samples were taken both in the P. australis and in Chara of the Glanas and Saby areas of the lake. The samples were sieved and taken in plastic buckets with a lid. In the laboratory, the benthic samples were sorted in a flat bowl containing clean water and the invertebrates were picked into glass cylindrical tubes and preserved in ethanol. The invertebrates were then identified into their various families with the aid of a microscope and an invertebrate manual. The length of the invertebrates was measured in (mm) with the aid of square paper mark placed under a Petri disc and observed through the microscope. Two similarity indexes were used to compare the proportion of invertebrates in the two habitats; these were Sorensen and Renkonen similarity index. While Renkonen consider the proportion, Sorensen uses the number of species as in the equation ii:
100
2
b
a
c
S
……...iiwhere c is the number of species that were found in both habitats, a is the number of species in location 1, b is the number of species in location 2.
Fish sampling was conducted in Lake Takern twice in autumn 2008 at Glanas and Saby area of the lake, using a standard survey gill net. The gill net was set both in the P. australis and in the Chara areas Data from another sampling occasion in autumn 2009 were also used. Gill nets were exposed to the water for two hours.
The fish were removed from the gill net, sorted, and the total length and weight were measured. Samples of the fish tissues were taken and subsamples of the fish were taken for gut content examination. Pair of surgical blade and scissors was used for this purpose; the stomach content were in turn introduced to Petri disc and then examined with the microscope to identify invertebrates’ parts remaining.
2.3 EXPERIMENTAL DESIGN
In the laboratory, perch predation on G.pulex and Corixidae (water boatman) of size 7mm, 9mm and 10mm was investigated using 100mm- 105mm size of perch. The perch were bought from the local fish farm, while the invertebrates prey were sampled in Lake Takern and ponds in Linkoping University. Three size groups each of G.pulex, Corixidae and A.
aquaticus were fed to 3 perch at different feeding occasions, in an aquarium fitted with
aerators and filters. The aquarium was partitioned with a vertical slide glass attached to a string which manually released to partition the fish and prey or pulled to remove the barrier, thus creating fish and prey access. There was also a paper screen to minimize human interaction with the fish activity in the aquarium. The fish activities in the aquarium were recorded by a camera mounted on a tripod. Three replicates were made using 3 different groups of perch. There were three holding plastic tubs in which the fish were first adapted to the prey items and also served for holding the fish from the aquarium.
9
The handling time was defined as the time between the attack of one prey and the predator’s readiness to pick another prey. These were estimated in each feeding occasion using a stop watch. The handling time observed from video by Andersson (2010), who was looking at the handling time for A.aquaticus in a similar experimental set up). In all, there were three different prey organisms and only one predator fish species. Ten individual prey were introduced to the aquarium stocked with perch in a group of three individuals at a time and one individual was observed and the handling time taken. The procedure was repeated 7 times for each of the prey (Corixidae, G.pulex and A.aquaticus) and their size categories using different fish individuals. 21 individual fish were used to estimate the average handling time. 2.4 DATA AND STATISTICAL ANALYSIS
Primary data were obtained both from the field sampling and the experimental set up. Data from the field include fish length and weight relationship, the benthic invertebrates’ distribution, fish gut content among others. The percentage distribution of the benthic invertebrates in the water as compared to the percentage found in the fish gut was estimated using a Chi square test. In the experimental set up, data on perch handling time for the three different invertebrate categories were obtained and recorded in an Excel work sheet. The mean, standard deviation and the covariance of the samples were calculated. The rate of biomass consumption by the predator was also estimated using length / body mass relationship sourced from literature (Haas et al.2007).
Table 1. Equations for body length (L) transformation to body dry weight (BDW) and calculation of handling rate
Handling rate = t wt time handling t body weigh …………... iii b L a BDW exp …... iv where a and b are constants taken from1,2 ;
L b a BDW 1 1 1 ………... v A. aquaticus1 : a 1 = -4212; b =2268... vi Corixidae1: a 1 = -4286; b = 2785... vii G. pulex2: a 1 = -4.783; b= 3.258... ... viii 1
Haas et al. 2007, 2 Meyer, 1989
Test of results were carried out using similarity indexes and test of paired proportions (t- test) for invertebrates distribution in the two contrasting habitats. Also, an ANOVA analysis for the mean handling time for perch predation on G.pulex, Corixidae and A.aquaticus was carried out.
10
3 RESULTS
3.1. FISH SAMPLES
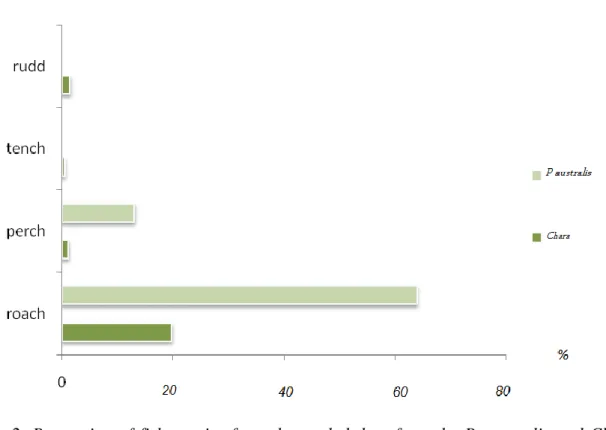
The samples of the fish taken from the gill net were mainly roach (Rutilus rutilus) and perch (Perca fluviatilis), whereas tench (Tinca tinca), and rudd (Scardinius erythrophthalmus) (Table 2) were only found in small numbers. The most common was roach and there were more fish in the P. australis vegetation (Fig. 2). The size of perch caught in the P. australis habitat was 98mm average and STD of 25, while in Chara perch size was 128mm average, with a STD of 36.
Table 2. Number of fish individuals sampled in Lake Takern at two occasions in 2008.
Date Location roach perch Tench rudd CPUE
Glänås 09/19/08 Chara 18 3 0 6 27 Säby P.austalis 54 1 0 0 55 09/24/08 Chara 67 2 2 0 71 P.australis 221 56 0 0 277
Figure 2. Proportion of fish species from the pooled data from the P. australis and Chara habitats in Lake Takern in 2008.
11 3.2 THE BENTHIC INVERTEBRATES SAMPLES
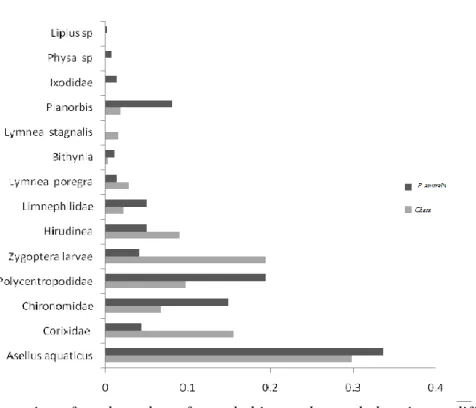
14 families of invertebrates were identified from the samples (Fig. 3). There were more invertebrate taxa represented in the P. australis than in the Chara habitat.
Figure 3. The proportion of total number of sampled invertebrates belonging to different taxa in Chara andP. australis habitats in Lake Takern in 2008.
The results from the calculated Renkonen and Sorensen similarity indexes show that the two habitats were similar in the distribution of invertebrate’s species the index was 65%.
Further, a paired t-test was made between the proportions of taxa in the two habitats and the results showed no significant differences between the frequency distribution (Table 3).
Table 3. Paired Samples t-test of the invertebrates sampled from Chara and P. australis habitats in lake Takern in 2008.
Paired differences
Chara invert freq. - P. australis invert freq.
Mean Std. dev Std. Error mean 95 % Confidence. interval of the difference t Df Sig. (2-tailed) -.00073 .06837 .01827 -.04020 .03874 -0.040 13 0.969
12 3.3FISH GUT CONTENT
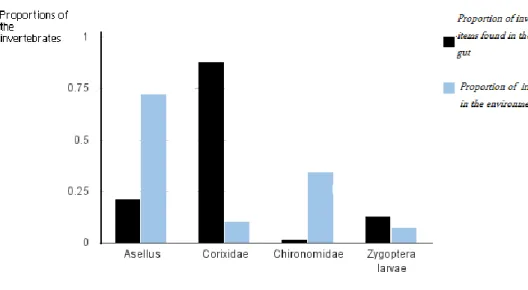
Four different invertebrates were found to be eaten by the perch (Fig.4).
Figure 4. The proportion of individuals among the four most frequently found invertebrates in the gut of Perch, the fish caught by the gillnet, and in the environment.
The chi square was estimated at 762 (Appendix 1), which was greater than the critical value of 7.8 at a degree of freedom 3 that is (n-1), n , number of individuals and at 0.05 error level. Thus the results indicate a significant difference between the distributions.
3.4 FISH PREDATION EXPERIMENT
In the fish predation experiment, there was no significant differences in the perch handling time for the different prey sizes, neither for Corixidae, G. pulex nor A. aquaticus (Fig. 5) and (Table 4-6). There was a significant difference in the intake rate for A.aquaticus, G.pulex and
13 Handling time S
A
5mm 7mm 9mmB
Handling time S 7mm 9mm 10mmC
Handling time S 7mm 9mm 10mmFigure 5. Perch predation on A) A.aquaticus, B) G. pulex, and C) Corixidae of three different sizes.
14
Table 4 Results from ANOVA analysis of experimental data on handling time and intake rate of perch predating on Corixidae
Sum of Squares df Mean Square F Sig. Intake rate mg/sec Between Groups .006 2 .003 .441 .663 Within Groups .042 6 .007 Total .048 8
Handling time Between Groups .434 2 .217 .219 .809
Within Groups 5.941 6 .990
Total 6.375 8
Table 5 Results from ANOVA analysis of experimental data on handling time and intake rate of perch predating on G.pulex
Sum of
Squares
df Mean
Square
F Sig.
Intake rate Between Groups 0.057 2 0.028 8.175 0.019
Within Groups 0.021 6 0.003 Total 0.078 8 Handling time Between Groups 1.267 2 0.633 2.253 0.186 Within Groups 1.686 6 0.281 Total 2.953 8
Table 6 Results from ANOVA analysis of experimental data on handling time and intake rate of perch predating on A.aqauticus
Sum of
Squares
df Mean
Square
F Sig.
Intake rate Between Groups .249 2 .125 13.548 .000
Within Groups .166 18 .009 Total .415 20 Handling time Between Groups .761 2 .381 1.196 .325 Within Groups 5.727 18 .318 Total 6.488 20
15
A
B
C
Intake rate mg/s Intake rate mg/s Intake rate mg/s 5mm 7mm 9mm 7mm 9mm 10mm 7mm 9mm 10mmFigure 6. Perch intake rate when feeding on A) A.aquaticus, B) G. pulex, and C Corixidae of three size classes.
16
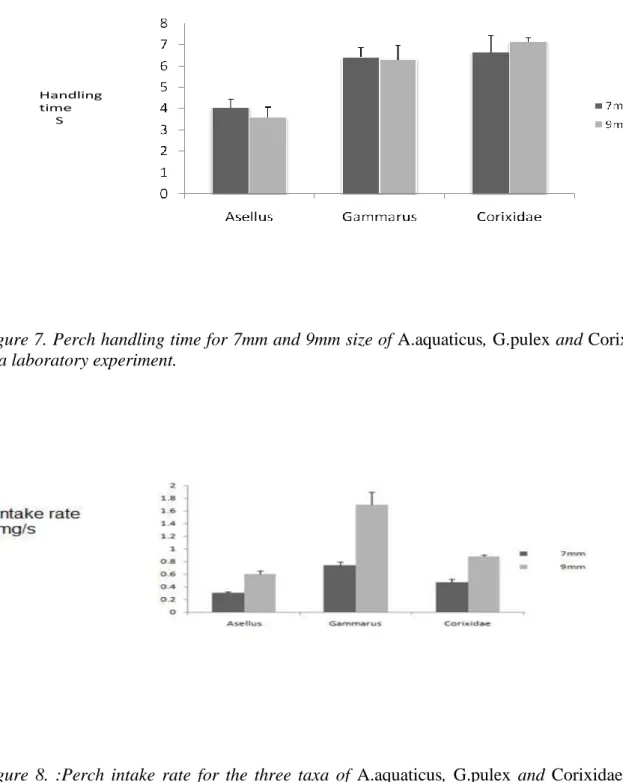
The data for the handling time and the respective intake rate of perch across the three prey taxa of sizes 7mm and 9mm were compared (Fig 7 and 8). The results from a two way ANOVA indicated that there were significant effects of both the species and their sizes on the intake rate (Table 7); and that the size of the prey contributed in the same way in the three species ( Species* Size p>0.05).
Handling time
S
Figure 7. Perch handling time for 7mm and 9mm size of A.aquaticus, G.pulex and Corixidae in a laboratory experiment.
Figure 8. :Perch intake rate for the three taxa of A.aquaticus, G.pulex and Corixidae in a laboratory experiment
17
Table 7: Results from a two way ANOVA of the effects of species and size of the prey on perch intake rate. Data were log-transformed in order to eliminate differences in variance.
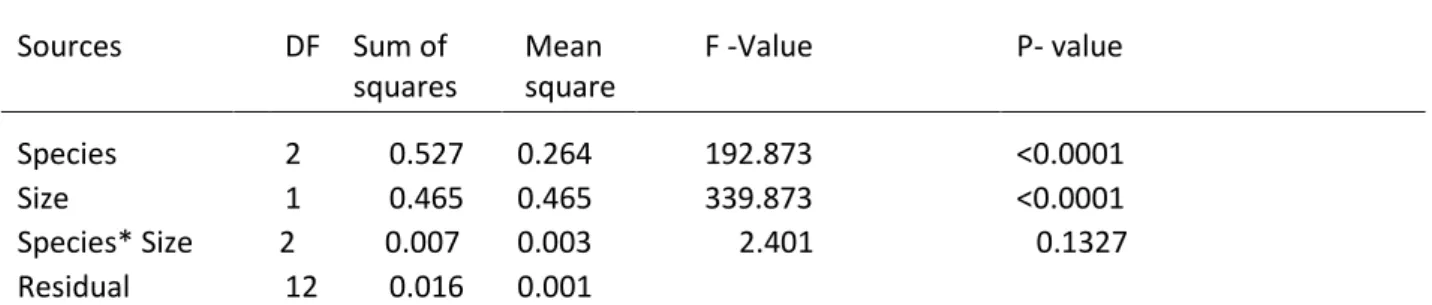
Sources DF Sum of squares Mean square F -Value P- value Species 2 0.527 0.264 192.873 <0.0001 Size 1 0.465 0.465 339.873 <0.0001 Species* Size 2 0.007 0.003 2.401 0.1327 Residual 12 0.016 0.001
18
4 DISCUSSION
4.1 SAMPLING TYPES
In the investigation of the research questions, sampling were carried out in the field and in the experimental set up. In the fish sampling the roach were more in number but their gut content analysis revealed so little of their prey organisms but for the perch it was possible to identify different prey organisms. Thus using perch as the benthic predator in the predator and prey experiment was an adequate choice.
Also, hand net samples of benthic invertebrates samples were taken in both P. australis and
Chara macrophytes vegetation area of the lake. However, there were no differences in the
benthic invertebrate taxa distribution; there was the general habitat complexity in the two areas.
Sorensen similarity index was calculated as 65%. This indicates that the similarity of the two habitats is above 65% by any standard procedure. Then what can one say about the dis- similarity, and the test of significance of such differences? The test of significant differences between pairs of frequencies using standard test such as a t- test shows p value of 0.969 which indicates no significant differences between the pairs.
4.2 FISH PREDATION ON INVERTEBRATES
The invertebrates’ distribution in an aquatic environment is a noted factor that affects the fish carrying capacity (Wetzel 2001). The fish depends on a wide variety of invertebrates as prey at different stages of their life cycle. Fish has been described as the dominant habitat regulator, a top predator in an aquatic system (Vanni 1986, Hansen and Jeppesen 1992, Scheffer 1998). The presence or absence of some species of invertebrates in a lake is used as an indicator of fish or fishless lakes in some recent work (Schilling et al. 2009). In the fish predation experiment, perch predation on A. aquaticus, G. pulex, and Corixidae was observed. There was evidence of the perch catching, chewing and swallowing the prey organism. Some handling time may be longer due to an incorporating search time in between actions. Handling time also include both the time for prey manipulation and the time the prey evacuate the stomach to the intestine (gastric evacuation), described as the total handling time (Nilsson and Bronmark 2000). Handling time may also be due to the prey size, the predator experience and the prey type (Bromley 1994, Andersen 1999). Preys with soft and small size body are likely to be taken more easily than those with rough body both in the manipulative stage and gastric evacuation stage.
In the experiment, the observed handling time defined as the manipulative period for A.
aquaticus was observed to be 4s at most, but 6s and 7s for G. pulex and Corixidae. One can
explain that the prey characteristics may have some impact on the predator handling time; G.
pulex is known to swim faster compare to A. aquaticus, while Corixidae are also fast
swimmers and with an adaptive rough body texture. Considering the gut content analysis,
Corixidae was more common as a prey item compared to other prey. This might also be a
factor of the nature of the body surface affecting the gastric evacuation time (Nilsson and Bronmark 2000). Also, Corixidae is often seen dangling between patches of submerged vegetation and the water surface, thus likely a more vulnerable prey to predators. A. aquaticus might be less vulnerable because of its association with the vegetation and the bottom substrates (Rooker et al 1998). This type of behaviour is likely a factor that can predict what types of prey items that will occupy the niche of the predatory fish species. The number of
19
perch individuals that were caught in the Chara vegetation was also more, which could be due to the favourable environment for predation on invertebrates and other small fishes.
4.3 RESULTS AND THE RESEARCH QUESTION
The research questions constructed from the hypotheses include: Are there any significant difference in the macro invertebrate species composition and their frequency in the two habitats? Secondly, are there any differences in the handling time for G. pulex and other invertebrates of the same size? The statistical evaluations of these two questions were as follows.
The benthic invertebrate distribution in the P. australis and Chara dominated areas showed no significant difference using a t- test, p>0.05. While the two habitats were similar, with a similarity index above 65%, there were some differences. The P. australis habitat had more taxa than the Chara habitat but the overlaps showed that the two habitats were indeed similar. The reason for the higher number of invertebrates taxa in the reed could be the complexity of the vegetation, that attract different types of invertebrate’s organisms. The Chara area is well dominated by Chara vegetation, which can be seen as a large cluster of Chara plants just underneath the water. Predation also is another source of community structuring. One can expect predation from fish to be more in Chara vegetation than in the P. australis because of the differences in their vegetative structures and physical characteristics which could affect the predator efficiency.
A. aquaticus is a common invertebrate prey organism in freshwater. In the experiment it was
preyed upon with the shortest handling time. It could be because of the soft body texture and the small size. The handling time is also related to the gastric evacuation rate (total handling time), which is directly related to the temperature, prey type and prey size. Perch had been reported to have a similar gastric evacuation rate across some invertebrates including: G.
pulex, Chaoborus, chironomids and zooplankton (Persson 1979, Singh-Renton and Bromley
2005).
Equally important is the comparison of the proportion of the prey items found in the gut of perch to the observed handling rate. The question is if the prey with higher proportion in the gut had less handling time or a higher intake rate (mg/sec). From the results of the perch gut analysis, Corixidae were the most common prey found in the fish gut but considering the handling rate, perch intake rate for G. pulex was higher than Corixidae. Thus, the proportion found in the gut may not necessarily mean that the particular prey organism has a proportional optimal foraging benefit for the predator. In another set of gut content analysis carried out in september 2009, Corixidae were also observed to be more common in the stomach of perch and the ratio Corixidae/A.aquaticus, was 4.2 ( unpublished data). The abundance of Corixidae in the stomach of perch also corresponded to the observed data I got in the stomach analysis.
20
4. SUMMARY AND CONCLUSION
The invertebrate distributions in the two habitats, P. australis and Chara submerged vegetation, were not significantly different.
In the predation experiment, perch predation on G. pulex of all sizes, Corixidae of all sizes and A.aquaticus of all sizes showed no significant differences in the handling time within the respective groups. Considering the intake rate of prey per unit time, the results showed that the time spent by predator fish across the three taxa was not significantly different, thus the intake rate (mg/s) increases with an increase in the prey size. This supports the optimum foraging theory that predators will preferably select prey items that is more beneficial. In all, the observed handling time shows that perch predation on A. aquaticus took the shortest handling time, followed by Corixidae and lastly G. pulex, but looking at the intake rates G.
pulex was a more optimal foraging prey for the perch than A aquaticus.
Considering the gut content analysis, perch had G. pulex, A. aquaticus and Corixidae in their gut. Corixidae were observed to be more common in the gut of perch. Therefore, A. aquaticus and G. pulex are just as likely a good prey organism for perch. It will be important to also consider the search time, because most handling time is followed by a time lapse for search planning of prey attack. In the natural water, where there is an abundance of prey refuges, the search time will be very important in a predator and prey situation. A.aquaticus and G. pulex are more likely to gain more protection from the submerged vegetation in the P. australis due to their constant use of the vegetation for both feeding on attached dead leaves and predator avoidance. Thus, perch predation on invertebrate would be higher in Chara habitat than in the
P. australis habitat due to the differences in the vegetation and turbidity of the two habitats.
Acknowledgment
I am sincerely grateful to the academic team, administrators and service personnel of the department of biology, Linköping’s University for their relentlessness and commitment to their work and service, thus paving way for the successful completion of the course work and project. More so, my greatest regard to my supervisor Anders Hargeby for his constructive criticism and objectivity; friends and family members for their generous emotional support all the time.
21
LIST OF REFERENCES
Andersen N.G (1999). The effects of predator size, temperature, and prey charactersitics on gastric evacuation in whiting. Journal of Fish Biology 54, 287-301.
Andersson M (2010). Selective predation by perch on freshwater isopod in two macrophyte habitat.
Video records of perch predation in laboratory experiment, MSc thesis
LITH-IFM-A-EX--10/2307-SE, Linkoping University.
Bechara J.A., Moreau. G and Hare. L.(1993). The impact of brook trout (Salvelinus fontinalis) on an experimental stream benthic community, the role of spatial and size refugia. Journal of Animal Ecology 62, 451- 464.
Beckett D. C., T. P. Aartila and A. C. Miller (1992). Invertebrate abundance on Potamogeton nodosus: effects of plant surface area and condition. Canadian. Journal of Zoology 70, 300-306.
Blumenshine S. C., Lodge D. M. and Hodgson J. R.(2000). Gradient of fish predation alters body size distributions of lake benthos. Ecology 81, 374–386.
Bromley P.J (1994). The role of gastric evacuation experiments in quantifying the feeding rates of predatory fish. Reviews in Fish Biology and Fisheries 4, 36- 66.
Bronmark C(1994). Effects of tench and perch on interactions in a freshwater, benthic food chain. Ecology 75, 1818-1828.
Charnov E. L (1976). Optimal foraging, attack strategy of mantid. American Naturalist 110, 141-151. Crowder L. B. and Cooper W. E (1982). Habitat structural complexity and the interaction between blue gills and their prey. Ecology 65, 894- 908.
Diehl S (1992). Fish predation and benthic community structure, the role of omnivory and habitat complexity. Ecology 5, 1646-1661.
Elliott J.M. (1976). The energetics of feeding, metabolism and growth of brown trout (Salmo trutta L) in relation to body weight, water temperature and ration size. Journal of Animal Ecology 45, 923-948 Elner R. W. and Hughes R.N (1978). Energy maximization in the diet of the shore crab, carcinus maenas . Journal of Animal Ecology. 47, 103-116.
Emlen L. M (1966). The role of time and energy in food preference. American Naturalist 100, 611-617.
Gilinsky E.(1984). The role of fish predation and spatial heterogeneity in determining benthic community structure. Ecology 65, 455-468.
Haas K., Kohler U., Diehl S., Kohler P., Dietrich S., Holler S., Jaensch A., M. Niedermaier M. and
Vilsmeier J (2007). Influence of fish on habitat choice of water birds: Ecology 88(11), 2915–2925.
Hansen A. M. & Jeppesen E(1992). Changes in the abundance and composition of cyclopoid copepods following fish manipulation in eutrophic lake, Denmark. Freshwater Biology 28, 183–193.
22
Hughes R.N. (1988). Optimal foraging in the intertidal environment, evidence and constraints. In Chelazzi, G., Vannini, M. (Eds). Behavioral adaptation to the intertidal life. NATO ASI Ser. A . Vol 151. Plenum press , New York, pp 265-282.
Hughes R. N., Seed. R (1995) Behavioral mechanism of prey selection in crabs. Journal of Experimental Marine Biology and Ecology 193, 225- 238.
Jackson A.C. & Rundle S. D (2008). Diet shifts by an estuarine goby (Potamoschistus microps) in the face of variable prey availability. Journal of Experimental Marine Biology and Ecology 361, 1-7. Jackson A.C., Rundle S. D, Attrill M.J and Cotton P.A (2004). Ontogenetic changes in metabolism may determine shift for a sit and wait predator. Journal of Animal Ecology 73, 536-545.
Johnstone I. and Norris K. (2000). Not all oystercatchers. Haematopus ostralegus, select the most profitable common cockles cerastoderma edule: A difference between feeding methods. Ardea 88. 137-153.
MacArthur R. H. and Pianka E. R. (1966). Optimal use of patchy environment. American Naturalist 100, 603- 609.
Meyer E.(1989). The relationship between body length parameters and dry mass in running water invertebrates. Archive Hydrobiologia 117, 191-203.
Mistri M. (2004). Predatory behavior and preference of a successful invader, the mud crab, Dyspanopeus sayi cpanopeidae, on its bivalve prey. Journal of Experimental Marine Biology and Ecology 312, 385-398.
Mittelbach G.G (1981). Foraging efficiency and body size, a study of optimal diet and habitat use by bluegills. Ecology 62, 1370- 1386.
Nilsson B. I.(1981). Susceptibility of some odonata larvae to fish predation. Limnologie 21. 1612-1615.
Nilsson P. A and Bronmark C (2000). The role of gastric evacuation rate in handling time of equal- mass rations of different prey sizes in northern pike. Journal of Fish Biology 57, 516- 524).
Persson L (1979). The effects of temperature and different food organisms on the rate of gastric evacuation in Perch (Perca fluviatilis), Freshwater Biology 9(2) 99 - 104
Persson, L. and Greenberg L. A (1990). Optimal foraging and habitat shift in perch (perca flaviatilus) in a resource gradient. Ecology 71. 1699- 1713.
Rooker J.R, Holt S.A, Soto M. A. and Holt G. J (1998). Post-settlement patterns of habitat use by sciaenid fishes in subtropical Seagrass meadows. Estuaries 21:315–324
Rutten A. L, Oosterbeck K., Ens B. J.and Verhulst S (2006). Optimal foraging on perilous prey; risk of bill damage reduce optimal prey size in oystercatchers.
Behavioural Ecology 17, 297-302
Scheffer M. (1998). Ecology of Shallow Lakes. Chapman and Hall. London. 357 pp.
Schilling E . G , Loftin C. S. and Huryn A. D (2009). Macroinvertebrates as indicators of fish absence in naturally fishless lakes. Freshwater Biology 54 (1) 181- 202.
23
Singh-Renton S. and Bromley P. J (2005). Effects of temperature, prey type and prey size on gastric
evacuation in small cod and Whiting:Journal of Fish Biology 2005; 49(4) 702 – 713
Smallegange M. I., Hidding B., Eppenga J. M..A and van der Meer J.(2008). Optimal foraging and risk of claw damage: How flexible are shore crabs in their prey size selectivity? Journal of Experimental Marine Biology and Ecology 367, 157-163.
Tolonen K. T., Hämäläinen H. Holopainen I . J, Mikkonen K., & Karjalainen J.(2003). Body size and substrate association of littoral insects in relation to vegetation structure. Hydrobiologia 499, 179-190. Vanni M. J. (1995).Fish predation and zooplankton demography. Ecology 67: 337–354.
Visser M. E.(1995).The effects of competition on oviposition decision of leptopilina heterotonia (Hymenoptera. Eucoilidae). Animal Behaviour 49, 1677- 1687.
Wellborn G.A, Skelly D. K. and Werner E.E.(1996). Mechanism creating community structure across a freshwater habitat gradient. Annual Review of Ecology and Systematics 27, 337-363.
24 Appendix
1 Estimating the invertebrates’ abundance in the benthic envioronment.
A aquaticus Corixidae Chironomidae Zygoptera larvae Benthic samples 88 13 42 9 Fish guts samples 26 106 2 16 Benthic prop 0.578947 0.085526 0.276316 0.059211 Fish guts proportion 0.173333 0.706667 0.013333 0.106667
A.aquaticus Corixidae Chironomidae Zygoptera larvae
Found in gut 26 106 2 16 n
Abundance in the benthic samples 86.84211 12.82895 41.44737 8.881579 150
X2=sum all (found -expected)2/expected
42.62635 676.6607 37.54388 5.705283
X2= 762.5362
II The intake rate of perch on its prey.
Size A.aquaticus G.pulex Corixidae
mm mg/sec mg/sec mg/sec
7 0,31 0,78 0,47 7 0,27 0,758 0,525 7 0,31 0,687 0,417 9 0,68 1,63 0,89 9 0,58 1,95 0,89 9 0,63 1,59 0,85