Bentiska evertebraters förekomst i källor
på Öland
Lina Hilmersson
Examensarbete i biologi
Nivå: D
Nr: 2009:Bi3
Examensarbeten gjorda vid Högskolan i Kalmar, Naturvetenskapliga institutionen, och lista över dessa kan beställas via; www.hik.se/student
eller: Högskolan i Kalmar Naturvetenskapliga institutionen 391 82 KALMAR Tel 0480-44 62 00 Fax 0480-44 73 05 e-post: info@nv.hik.se
Examensarbeten gjorda på Högskolan i Kalmar finns på: www.hik.se/student
Bentiska evertebraters förekomst i källor på Öland
Lina Hilmersson
Biologiprogrammet 160 p, Högskolan i Kalmar, Naturvetenskapliga Institutionen
Examensarbete, (Biologi): 30 hp för Filosofie Magisterexamen
Handledare: Docent Jan Herrmann Högskolan i Kalmar
Examinator: Universitetslektor Geoffrey Lemdahl Högskolan i Kalmar
Abstract
The benthic macroinvertebrate fauna of 22 natural springs on Öland was sampled, by using a time-standarized kick/sweep sampling technique, in the end of September and early October. The aim was to determine the number of species and their abundances in the macroinvertebrate communities in springs and how/why these may change between different springs. Likewise there was a purpose to investigate possible differences between springs considering the proportions of the number of species and the abundances of the five functional feeding groups; shredders, collectors, filter-feeders, scrapers and predators. There was also an aim to investigate if the distances betweeen springs had any influence on similarity between the communities of benthic macroinvertebrates of the springs. Analysis from the springs collected material showed several interesting patterns in the number of species and their abundances when compared between springs devided into groups depending on their environmental factors such as depth and pH. Equally interesting patterns where found in the proportions of the number of species and their abundances of the functional feeding groups between different groups of springs with different environmental features such as size, depth and type of surrounding area. Results also showed that springs separated by shorter distances had a higher likeness between their communities of benthic macroinvertebrates then springs separated by greater distances.
Innehållsförteckning 1. Introduktion
¾ 1.1 Källan s. 3
¾ 1.2 Källvattnets kemiska egenskaper s. 3
¾ 1.3 Källtyper s. 4
¾ 1.4 Livet i och runt källan s. 4
¾ 1.5 Syfte s. 5
2. Material och metoder
¾ 2.1 Metodik s. 6
¾ 2.2 Analys i Primer 6.1.2. s. 7
3. Resultat
¾ 3.1 Vattenegenskaper och källstorlek s. 8
¾ 3.2 Bottensubstrat, detritus och död ved s. 8
¾ 3.3 Närmiljö och vegetation s. 8
¾ 3.4 Bottenfauna s. 9
¾ 3.5 Resultat av analys i Primer s. 14
¾ 3.6 Funktionella grupper s. 17
4. Diskussion s. 17
5. Sammanfattning s. 19
6. Tack s. 19
7. Referenser s. 20
Bilaga 1 - Lokalbeskrivningar, vattenkemi, bottensubstrat, markanvändning,
närmiljö och vegetation s. 22
1. Introduktion
Öland är känt som Sveriges minsta landskap men även som ett av de mest intressanta för sina natur- och kulturaspekter. Ön omtalas ofta som mångfaldens ö bl.a. för att den ger möjlighet att beskåda naturen ur en mängd olika vinklar, här kan man uppleva allt från vindpiskade klippor till gamla lundar. Det här arbetet är viktigt, för att det undersöker en ny biologisk aspekt av den Öländska naturen, nämligen källorna, som hittills inte uppmärksammats i ett landskap som så ofta hyllas av naturälskare.
1.1 Källan
En källa är enligt definitionen ett distinkt (från en punkt) utflöde av grundvatten ur berg eller jord samt den ansamling av vatten med tillhörande avrinning som följder ett sådant utflöde. Vad som avgränsar en källa från en annan kan vara svårt att bestämma. I ett och samma område med liknande geologi och topografi kan flera utflöden finnas och beroende på hur snävt man avgränsar definitionen på en källa, kan dessa utflöden räknas som separata källor eller en och samma källa med mer än ett utflöde. Utflödet behöver heller inte leda ut från jord och berg till atmosfären utan kan även mynna under vatten t.ex. i sjöar och i havet. (Källakademins redaktionskommite, 2006). Ur ett biologiskt perspektiv är källorna också intressanta, då de kan erbjuda en ovanlig miljö med bl.a. relativt konstant temperatur och flöde (Bråvander m.fl. 2006). Källor är viktiga i naturen eftersom de dels kan utgöra en vattenkälla för större djur under vintern då andra vatten fryser men även eftersom de tillsammans med andra inslag i naturen, så som bäckar, dungar och ängar mm, bidrar till den biologiska mångfalden. Enligt en inventering av olika vattenmiljöer i södra Hälsingland, utförd av Per-Ola Hoffsten med kollegor, bidrog källorna med 10 % av den biologiska mångfalden vilket kan ge en fingervisning om källornas betydelse. Källor utgör även en sista utpost för reliktarter från en tid då Sverige bestod av en tundramiljö och kan förklaras som nyckelbiotoper om nationellt hotade arter förekommer i eller runt källan (Hoffsten 2006). 1.2 Källvattnets kemiska egenskaper
Källvattnets kemiska egenskaper påverkas av bl.a. vattnets uppehållstid i marken och vilken typ av jordlager/berggrund vattnet infiltreras genom. Dessa faktorer har stor inverkan på vilken kvalitet och vilka egenskaper källans vatten får. Den nederbörd som når marken förändras kemiskt sett på vägen ner mot grundvattnet, bl.a. av atmosfärisk koldioxid. I det övre marklagret kommer den mikrobiologiska verksamheten sedan att konsumera syre samt bilda koldioxid vid nedbrytning av organiskt material (Hult 2006). När koldioxiden löses i
vattnet bildas i en kemisk process kolsyra som bl.a. sänker pH (H2O + CO2 ⇌ H2CO3)
(Oxford University Press 2004). Vattnet löser även ut humusämnen och mineraler, något som påskyndas av ett lägre pH-värde. Lösta mineraler kan sedan fällas ut längre ner i marken och bilda s.k. rostjordar. Förekomsten av aeroba bakterier och virus som finns i de övre jordlagren minskar i takt med att syrehalten sjunker desto längre ner i marken vattnet infiltreras. Även förekomsten av organiskt material nedbrutet i olika utsträckning samt humusämnen kommer att minska då de sakta filtreras bort i marken. Hur väl vattnet filtreras beror även på typen av mark det rinner genom samt uppehållstiden. Vatten som filtreras genom finare sediment t.ex. sand eller lera är ofta renare än det som runnit genom större sprickor i berggrund (Hult 2006).
mineraler och salter vattnet innehåller d.v.s. hur mycket av dessa vattnet hinner lösa ur marken och berggrunden, innan det återigen når markytan (Hult 2006).
Källornas vatten syresätts igen då det rinner upp i källutflödet, något som kan leda till att kemiska substanser fälls ut i större utsträckning. Sådana utfällningar kan till exempel vara kalciumkarbonat som med tiden kan omvandlas till kalktuff (Habdija 2001), något som ofta kan ses vid källor med kalkhaltigt vatten. Röda utfällningar av järn finns i form av järnockra (en blandning av järnoxider), något som, om det fälls ut i tillräcklig utsträckning, kan brytas för industriell användning (då den innehåller 62 viktsprocent järn) (Hult 2006, Wikipedia 2007). Vid anaeroba förhållanden kan svavelhaltiga mineral löses ut i vattnet och bilda svavelväte, en kemisk substans vars lukt påminner om ruttna ägg. Lukten försvinner dock om vattnet får reagera med syre och även om ämnet är giftigt finns det inte i tillräcklig mängd i Sveriges grundvatten för att det skulle kunna utgöra någon hälsorisk. Vid utflödet kommer källans vatten återigen, precis som när det föll som nederbörd och nådde marken, att påverkas av källans mikroorganismer och humusämnen samt markanvändningen runt om källan (Hult 2006).
1.3 Källtyper
Svenska källor brukar klassificeras efter geologi, topografi, framträngande och tryckförhållanden samt vattnets kemiska sammansättning. I Sverige finns åtta olika sorters källtyper, moränkällor, åskällor, svallgruskällor, artesiska källor, källkupoler, bergkällor, karstkällor och mineralkällor (Knutsson 2006b). Eftersom de källor som kommer att undersökas i detta arbete förväntas domineras av karstkällor följer här en kort beskrivning av denna källtyp.
Karstkällor – Karstkällor uppkommer i kalkberggrund där erosion och kemisk vittring har bildat sprickor och kanaler genom vilka grundvatten kan ta sig upp till ytan. Flödena i dessa källor kan vara måttliga med endas ett par liter per sekund som rinner fram, men inom denna typ av källor kan man hitta några av de största flödena där 1000-tals l/s forsar fram under spektakulära former. Precis som berg- och moränkällor saknas ofta mäktiga jordlager varpå vattenkvaliteten inte kan klassas som den bästa. På grund av det höga kalkinnehållet har vattnet ofta god buffringsförmåga och högt pH, utfällningar av kalktuff är vanligt. På Öland varierar ofta vattenflödet med nederbörden eftersom vattenmagasinen här är mindre. Den höga kalkhalten lockar ofta kalkälskande arter, såsom orkidéer, att växa runt dessa källor (Knutsson 2006b).
1.4 Livet i och runt källan
Smådjursfaunan i källor brukar domineras av arter som är krenofila, dvs. arter som är källälskande men som även förekommer i andra typer av miljöer. Den krenofila faunan är därför inte unik men likväl anpassad till källmiljön och uppgår ofta till över hälften av arterna. En mindre del av faunan utgörs av krenobionter vilka är arter, som till skillnad från krenofilerna, är anpassade endast till att leva i källor. Källor är även lämpliga miljöer för sådana arter som annars ofta hittas i mindre vattenförekomster, så som rännilar eller mindre vattensamlingar i anslutning till större, eftersom sådana miljöer ofta är mycket lika de förhållanden som råder i källans kantzon som ofta har en högre vattentemperatur än källans utflöde (Hoffsten 2006).
Eftersom källans vatten håller en temperatur något över noll och därmed sällan eller aldrig fryser under vintern, har många av dess smådjursarter långsam tillväxt året runt. Det vanligaste bland vattenorganismer är annars att växa snabbare då vattentemperaturen höjs och sedan tillbringa kallare perioder med nedsatt tillväxt. Trots att källor ger tillgång till en unik miljö med tämligen konstant flöde och temperatur, är de ofta relativt artfattiga i Sverige. Den svenska smådjursfaunan har inte lika många krenofila och krenobionta arter i jämförelse med södra Europa. Anledningen till detta tros vara att den södra delen av Europa inte påverkades av inlandsisen. De krenobionter som finns i våra källor idag räknas som relikter som blev kvar under skiftet mellan varma och kallare perioder. Vissa av dessa arter är även bundna till källor med kallt vatten och låga sommartemperaturer, något som var vanligt efter inlandsisens avsmältning och då landet dominerades av tundramiljö (Hoffsten 2006).
Den vanligaste funktionella gruppen i källor är fragmenterare som tar hand om det döda organiska växtmaterial som tillförs från omgivningen men som även produceras i källan. De vanligaste arterna brukar återfinnas inom ordningarna nattsländor, bäcksländor, tvåvingar (fam. Tipula) och likfotingar (fam. Isopoda), vars larver i ett första led finfördelar det större materialet såsom löv eller dylikt. Även så som de rester som blir kvar efter finfördelningen kan sedan fångas upp av findetritusätare såsom fjädermygglarver. Mindre partiklar kan även föras vidare med vattnet och då konsumeras musslor och knottlarver (filtrerare) som filtrerar vattnet. De alger som växer på bl.a. löv och stenar kan tas omhand av skrapare (ex snäckor) eller ibland någon av de andra grupperna när deras primära födokälla sinar. Predatorarter såsom flertalet skalbaggsarter livnär sig på bytesdjuren i källan men utgör i sin tur tillsammans med de andra smådjuren en födokälla för större rovdjur såsom groddjur och fåglar (Hoffsten 2006).
Källor kan utgöra refugier för växter i landskapet eftersom de erbjuder en miljö med ibland rörliga, syrerika och kalla vattenförhållanden som rinner upp i en miljö som ofta har konstant temperatur, ljustillgång och fuktighet. Trots sin stabilitet finns inga obligata källväxter i Sverige, däremot finns det några som är källälskande. Dessa arter (flertalet mossor) är anpassade till en speciell vattenkemi eftersom källors vatten ibland innehåller höga halter av t.ex. järn eller kalcium beroende på de hydrogeologiska egenskaperna i grundvattenmagasinet som föder källan. Floran behöver dock inte vara speciellt anpassad till källmiljön, det kan helt enkelt vara så att den trivs vid källan eftersom den erbjuder en fuktig miljö. En av faktorerna som styr hur floran ser ut runt en källa är hur ytligt grundvattnet ligger. Vatten som passerar genom mäktigare marklager har ofta högre pH och högre koncentration lösta joner än vatten från ytliga grundvattenförekomster. Hur stora fluktuationerna är i pH betyder mycket för hur floran ser ut eftersom de flesta växter är känsliga för större förändringar. Källor på Öland har ofta ytliga grundvattenförekomster men eftersom den underliggande berggrunden till största delen består av kalksten, har dessa källor vatten ofta ett konstant högt pH-värde. I kalkrika miljöer trivs bl.a. orkidéer och andra kalkälskande arter i och runt källan, men i de allra kalkrikaste miljöerna, där ofta kalktuff och bleke fälls ut, trivs endast ett färre antal arter (Bråvander et. al. 2006).
1.5 Syfte
källor på ett sätt som ger ett insamlat material präglat av så många av källans olika miljöer som möjligt. Ett fältprotokoll kommer även att uppföras för varje källa och en insamling av dominant vegetation utföras för att ge en beskrivning av källmiljön och området runtomkring med avseende på t.ex. markanvändning, vegetation och vattenegenskaper. Den insamlade informationen om källornas miljöer avses sedan användas för att om möjligt koppla ihop eventuella skillnader och/eller likheter i källornas evertebratsamhälle med tänkbara bakomliggande faktorer. Vidare kommer det även att undersökas om och i så fall hur avstånden mellan källorna påverkar likheten mellan olika källors evertebratsamhälle.
2. Material och metoder
2.1 Metodik
Eftersom kunskapen om källors placering på Öland inte finns samlad i någon gemensam litteratur var det nödvändigt att genomföra en undersökning om var tänkbara lokaler med källor kunde finnas. Genom studier av kartor, Sveriges Geologiska Undersöknings källarkiv, sökning via Internet, insamlad information från ett antal kontaktpersoner samt studier av relevant litteratur kunde ett större antal bekräftade källor, samt vattensamlingar som misstänktes ha ett källflöde, lokaliseras. Under slutet av september och början av oktober besöktes 25 lokaler med sammanlagt 36 källor. Av dessa var det 22 källor som kunde lokaliseras, och som ansågs lämpliga för håvning d.v.s. källor som inte var helt överväxta av vegetation, sinade eller med pumputrustning monterad i källflödet. Några källor som lokaliserats och som ansågs lämpliga för håvning föll dock ändå bort p.g.a. tidsbrist.
Vid varje källa samlades växter in i plastpåsar för förvaring och senare nyckling. Medtaget fältprotokoll (Naturvårdsverket 2001, Bilaga 2) fylldes sedan i med avseende på närmiljö, bottensubstrat etc. Djup- och längdmätningar utfördes så långt detta var möjligt och noterades i fältprotokollet. En skiss tecknades över lokalen med inritad norrpil samt symboler för de sedan tagna fotografiernas riktningar. Håvning skedde under 2 min 30 s eller 1 min 15 s beroende på den källika miljöns storlek. Arbetet utfördes på sådant vis att största möjliga del av källans mikrohabitat täcktes in av håvningen, under vilken bottensubstratet även rördes upp för att få med dess ingående fauna samt organismer som kunde tänkas försöka undkomma håven. Det insamlade materialet överfördes till 2-liters plastburkar med lock och spritades med 96 % etanol. Ett vattenprov togs före håvning vid det som sågs som eller misstänktes vara källans uppflöde. Konduktivitet, pH och temperatur mättes i tagna vattenprov med en Universal meter Multiline p4 (F/SET-3) producerad av WTW, placerad i det fordon som användes, då håvning etc. slutförts.
För att inte besväras av etanolångor eller partiklar som kunde försvåra grovsorteringen av den insamlade materialet sköljdes provet i en sil med en maskvidd 0,5 mm. Det sköljda provet placerades sedan i en större vanna med vatten ur vilken faunan sedan sorterades ut. I de fall det var möjligt användes subsampling för individgrupper som förekom i ett så stort antal att utplockning av varenda individ inte var tidsmässigt möjlig. Subsamplingen har antingen grundat sig på fem likstora fält i en utvald vannas botten där djuren i ett fält räknades och sedan multiplicerades med antalet fält för att få en uppskattning av det totala antalet djur, eller som en uppskattning av andelen prov, om individantalet per provburk till synes varit relativt jämnt. De utplockade djuren placerades sedan i mindre burkar med etanol för kommande nyckling.
Faunan bestämdes till art eller så lång det var möjligt med hänsyn till tillgänglig litteratur. Utöver identifieringen av faunan räknades även antalet individer inom respektive taxa vilka även indelades under korrekt funktionell grupp.
För att kunna klargöra vilka faktorer som förklarar eventuella skillnader i abundans och artantal mellan källorna, utfördes en gruppering av källorna med hjälp av insamlade uppgifter om pH, konduktivitet, djup, omkrets, närmiljö, andel vattenväxter, bete, beskuggning, bottensubstrat och vattenhastighet. Eftersom det inte finns några tidigare studier över dessa faktorer i Öländska källor gjordes indelningen utefter vad som kunde tänkas vara rimligt efter observationer i fält samt med hänsyn till tidigare kunskap om andra typer av vattenmiljöer. pH – högre/lägre, klasserna sattes till högre och lägre eftersom ingen uppfattning fanns om vad som är högt eller lågt pH i just källor, pH lika med eller över 7.5 räknades som högre (9 lokaler) och värden därunder som lägre (13 lokaler).
Konduktivitet – högre/lägre, även här sattes klasserna som högre och lägre av samma anledning som för pH, konduktivitet lika med eller högre än 350 räknades som högre (17 lokaler) och värden därunder som lägre (5 lokaler).
Djup – grund/djup, vid gruppering efter djup användes både medeldjup och maxdjup, källor med ett medeldjup under 0.4 m klassades som grunda (11 lokaler) men kunde klassas som djupa om maxdjupet skiljde sig mycket från medeldjupet. Källor med ett medeldjup större än eller lika med 0.4 m klassades som djupa (11 lokaler).
Omkrets – liten/mellan/stor, källor med en omkrets mindre än 20 m klassades som små (11 lokaler) medan de med en omkrets över 50 m klassades som stora (3 lokaler), källor med en omkrets över 20 m men under 50 m klassades som källor av mellan storlek (8 lokaler). Närmiljö – alvar/skog/gräsmark, baserat på miljön runt källan, alvar 8 lokaler, skog 7 lokaler och gräsmark 7 lokaler. Alvarmark har här räknats som ett tunt jordmånstäcke ovanpå kalkstenshäll som ibland går i dagen med sparsam vegetation med stort inslag av lavar, till skillnad från gräsmark som ofta är en av boskap betad miljö med tjockare jordmån och en vegetation mest bestående av gräs och örter,
Andelen vattenväxter – inga/lite/mellan/mycket, baserat på en skala från 0-3 vilket noterats i fältprotokollet, 0-inga (3 lokaler), 1-lite (10 lokaler), 2-mellan (5 lokaler) och 3-mycket (4 lokaler).
Övriga faktorer så som bete, beskuggning, bottensubstrat och vattenhastighet hade i likhet med andelen vattenväxter bedömts utefter en skala från 0-3 där 0 var inga och 3 var mycket. 2.2 Analys i Primer 6.1.2
För statistisk analys av de antal arter och deras ingående individantal, som tidigare nycklats och räknats, användes Primer 6.1.2. Primer är ett dataprogram utvecklat för icke-parametriska multivariatanalyser och signifikanstester av dessa. Analyserna kan t.ex. tillämpas på biologiska data så som djur- och växtsamhällen. Denna typ av data kräver ofta analyser som kan hantera flera variabler och som inte kräver att dataunderlaget följer någon typ av fördelning (t.ex. normalfördelning), detta tillfredställs i det valda programmet.
3. Resultat
En kortfattad beskrivning av de källor som använts för provtagning och deras närmiljö finns i Bilaga 1.
3.1 Vattenegenskaper och källstorlek
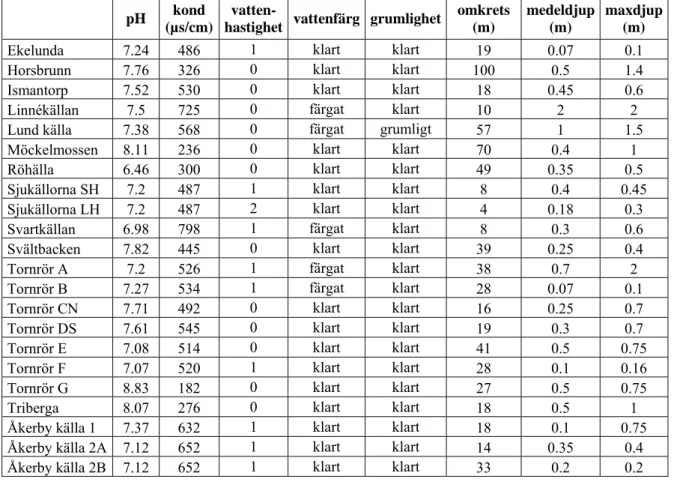
Vattnets pH i de olika källorna varierade mellan 6.46 (Röhälla, Bilaga 1, Tabell 1) och 8.83 (Tornrör G, Bilaga 1, Tabell 1). Konduktiviteten var högst i Svartkällan (798 µs/cm, Bilaga 1, Tabell 1) och lägst i källan Tornrör G (182 µs/cm, Bilaga 1, Tabell 1). Eftersom temperaturmätningarna gjordes på provtaget vatten och inte direkt vid källuppflödet avspeglade de troligtvis lufttemperaturen mer än källvattentemperaturen under provtagningsdagarna, beroende på hur lång transport-sträckan från källan till mätutrustningen var, och redovisas därför inte. Vattenhastigheten beskriver hur fort vattnet rör sig vid uppflödet då detta kunnat lokaliseras. Då uppflödespunkten inte varit möjlig att finna, har vattenhastigheten bedömts vara noll eftersom det då handlat om källdammar utan rinnande vatten. Vattnet i källorna har i alla fall utom ett (Lund källa, Bilaga 1, Tabell 1) varit klart, men ibland med färg (ofta grön- eller blåaktigt). Störst var källan Horsbrunn (100 m i omkrets, Bilaga 1, Tabell 1) och minst en av Sjukällorna (ca 4 m, en del av källflödet inräknat), storleken för de andra källorna har varierat däremellan. Djupet har varierat från några centimeter till över en meter, djupast var Tornrör A och Linnékällan vars maxdjup låg runt 2 m och kunde inte mätas fullt ut. Många av källorna har varit relativt grunda i kanterna men ofta med plötsliga djupskillnader längre ut.
3.2 Bottensubstrat, detritus och död ved
I de flesta källor har botten bestått av häll eller större stenar med varierande grad av fin- och grovdetritus (Bilaga 1, Tabell 2). Uppgifter om Linnékällans och Åkerby källa 1’s bottensubstrat saknas då detta inte kunnats bestämmas bland annat p.g.a. djup och siktsvårigheter. Istället har en uppskattning gjorts med källans utformning och omkringliggande miljö i åtanke. Mängden död ved har varit mycket sparsam eller saknats helt, även i källor med träd vid källkanten.
3.3 Närmiljö och vegetation
Marken runt källorna utnyttjas oftast som betesmark för hästar, kor och får, med olika grad av påverkan av källorna. Med närmiljö menas det område som finns från källkanten och 30 m utåt i alla riktningar. Närmiljön har i de flesta fall bestått av en blandning av alvarmark, gräsmark och trädbevuxna ytor med undantag av några källor, så som Horsbrunn (endast skog, Bilaga 1, Tabell 3) och Möckelmossen (endast Alvar, Bilaga 1, Tabell 3). Träd och buskvegetationen domineras ofta av björk, en och hassel med inslag av andra arter så som oxel och slån. Beskuggningen varierar mellan källorna beroende på hur nära och i vilken omfattning träd och buskar finns. Ett antal källor finns som inte skuggas alls eller delvis och endast tre (Horsbrunn, Svartkällan och Tornrör F, Bilaga 1, Tabell 3) har skog runt hela källan och skuggas därmed mer än de övriga.
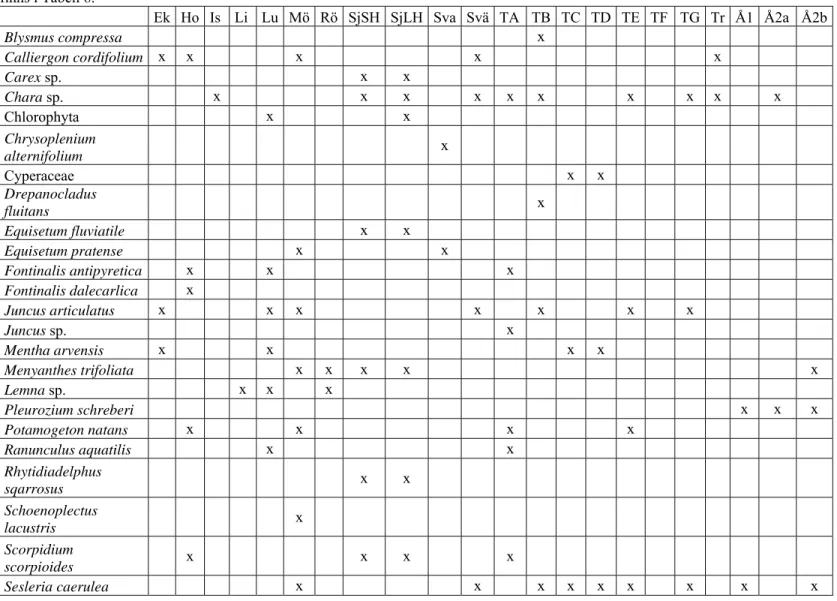
Övervattenväxter, flytbladsväxter, långskottsväxter och mossor dominerar i olika utsträckning källornas vattenvegetation som ofta är sparsam och ibland obefintlig (ex Tornrör F, Bilaga 1, Tabell 5). I Bilaga 1, Tabell 6 visas förekomsten av de dominerande växt- och mossarterna
(samt en algart Chara sp.) i och runt källan. Ryltåg (Juncus articulatus) och älväxing (Sesleria caerulea) är vanliga inslag runt källorna tillsammans med kransalger av släktet
Chara samt bla gäddnate (Potamogeton natans) som ofta förekommer i källorna.
3.4 Bottenfauna
Totalt för alla källorna hittades 79 taxa evertebrater (Tabell 7). Coleoptera var den största ordningen med 42 taxa, individrikast var dock ordningen Isopoda med en total abundans på 9213 individer fördelat på 2 taxa. De taxa som utmärkte sig från de övriga med fler än 1000 individer totalt på de lokaler de förekom i var Lymnea peregra och Planorbis planorbis (klass Gastropoda, 1941 resp. 3023 individer), Cloeon dipterum/inscriptum (ord. Ephemeroptera, 2829 individer), Corixa sp. (ord. Heteroptera, 1251 individer) samt Asellus aquaticus och
Gammarus pulex (ord. Isopoda, 7268 resp. 1585 individer). Flest arter (34) hittades i Tornrör
Tabell 7. Abundans, artantal och funktionell gruppering av bottenfauna. Abundansen är omräknad till provtagningstiden 2 min 30 sek. Förklaring till förkortningar av funktionella grupper kan ses i Tabell 8.
Ek Ho Is Li Lu Mö Rö SjSH SjLH Sva Svä TA TB TC TD TE TF TG Tr Å1 Å2a Å2b
Hirudinea
Funk. grupp
Glossiphonia complanata Pre 0 0 0 0 0 2 0 0 0 0 0 0 0 0 0 4 0 0 0 0 0 0
Haemopis sanguisuga Pre 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Herpobdella octoculata Pre 0 0 0 0 0 3 0 0 0 0 0 0 0 0 0 1 0 0 6 0 0 0
Gastropoda
Bathyomphalus contortus Fin 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 100 0 0 0 0
Bithynia tentaculata Skr 0 0 0 0 0 436 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Lymnea palustris Skr 0 0 0 0 0 0 0 0 0 0 10 7 0 61 54 0 0 380 0 0 0 0
Lymnea peregra Skr 6 0 2 0 0 1470 0 0 0 0 0 0 177 2 3 1 0 30 0 118 62 70
Lymnea stagnalis Skr 0 0 0 0 0 0 0 0 2 0 0 0 0 0 0 0 0 0 0 0 0 0
Physa fontinalis Skr 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 20 0 0 0 0
Planorbis planorbis Fin 0 3 44 0 0 415 0 0 0 0 1 0 0 0 0 107 0 2430 0 7 10 6
Bivalvia Pisidum spp. Fil 0 3 0 0 0 0 0 2 0 83 1 0 68 0 0 0 0 0 0 2 2 0 Acarina Hydracarina sp. Pre 0 0 0 0 0 36 0 0 0 0 0 0 0 0 0 0 0 0 18 0 0 0 Ephemeroptera Baetidae Skr 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 Caenis sp. Skr 0 0 0 0 0 2 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Cloeon dipterum/inscriptum Skr 0 56 256 0 58 0 42 0 0 0 1989 0 0 98 21 16 0 106 159 0 0 28 Odonata
Aeshna grandis Pre 0 2 2 0 0 0 0 0 0 0 2 1 0 0 0 77 0 0 0 1 2 0
Coenagrion lunulatum Pre 0 139 0 0 0 7 0 0 0 0 1 1 0 0 0 10 0 2 0 0 0 0
Ek Ho Is Li Lu Mö Rö SjSH SjLH Sva Svä TA TB TC TD TE TF TG Tr Å1 Å2a Å2b Odonata
Orthetrum cancellatum Pre 0 2 0 0 0 2 0 0 0 0 0 0 0 16 0 3 0 0 24 0 0 4
Heteroptera
Corixa sp. Pre 0 46 2 0 12 251 215 2 0 0 117 3 1 19 45 3 0 248 77 32 12 166
Gerris lacustris Pre 0 3 0 0 0 0 0 2 0 0 0 0 0 0 0 6 0 0 0 0 0 0
Nepa cinerea Pre 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 2 0 0
Notonecta glauca Pre 0 3 0 0 0 0 1 0 0 0 10 2 0 0 0 0 0 3 0 0 2 2
Notonecta maculata Pre 0 0 0 0 0 2 0 0 0 0 0 4 0 2 0 0 0 1 2 0 2 0
Notonecta obliqua Pre 0 0 0 0 0 0 0 0 0 0 0 3 0 0 0 0 0 0 0 0 0 0
Veliidae Pre 0 0 0 0 0 0 0 0 0 2 0 0 0 0 0 0 0 0 0 0 0 0
Coleoptera
Acilius canaliculatus Pre 0 1 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0
Acilius sulcatus Pre 0 3 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Agabus bipustulatus Pre 2 0 4 0 0 0 0 26 0 5 1 0 0 8 1 6 3 1 3 0 4 0
Agabus bipustulatus (larv) Pre 0 0 1 2 9 0 3 0 0 1 0 0 2 0 0 1 0 2 50 1 2 4
Anacaena lutescens Pre 0 1 5 0 0 0 0 0 0 0 0 0 0 0 0 0 0 7 0 0 0 0
Bidessus unistriatus Pre 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 2 0 0 0
Colymbetes fuscus Pre 0 0 0 0 0 0 0 0 0 0 2 0 0 0 0 0 0 1 0 0 0 0
Dytiscus marginalis Pre 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 2 0
Dytiscus semisulcatus Pre 0 3 5 0 0 0 0 4 0 0 0 0 0 5 4 17 1 0 1 0 0 0
Dytiscus semisulcatus (larv) Pre 0 0 10 0 0 0 0 0 0 0 0 2 0 0 0 0 0 0 0 0 0 0
Elmis sp. (larv) Fin 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0
Enochrus quadripunctatus Fin/Skr 0 0 1 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0
Enochrus testaceus Fin/Skr 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 14 0
Graptodytis granularis Pre 0 5 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Ek Ho Is Li Lu Mö Rö SjSH SjLH Sva Svä TA TB TC TD TE TF TG Tr Å1 Å2a Å2b Coleoptera
Haliplus obliquus (larv) Fin/Skr 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Haliplus ruficollis Fin/Skr 0 5 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 2 0 0 0 0
Haliplus ruficollis (larv) Fin/Skr 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 2 0 0 0 0
Helophorus grandis Fin/Skr 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 4 0
Helophorus granularis Fin/Skr 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Helophorus griseus Fin/Skr 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0
Helophorus minutus Fin/Skr 2 0 0 10 0 1 0 0 0 0 0 0 2 12 12 0 0 10 2 0 12 8
Hydrobius fuscipes Fin/Skr 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 2 0 0 0 0 0 0
Hydroporus erytrocepharus Fin/Skr 0 7 9 0 0 0 0 2 0 0 1 0 0 10 21 0 0 4 26 0 0 0
Hydroporus
palustris/incognitus Fin/Skr 0 17 0 3 1 0 0 10 0 0 0 0 0 35 12 0 0 0 14 0 0 0
Hydroporus planus Fin/Skr 0 0 1 18 0 0 0 2 0 0 0 0 0 0 0 0 0 0 1 0 0 0
Hydroporus pubescens Fin/Skr 2 0 0 4 0 0 0 0 0 0 0 0 12 0 0 13 0 4 55 0 2 0
Hygrotus confluens Pre 0 0 0 6 0 0 0 0 0 0 0 0 0 1 0 0 0 0 2 0 0 0
Hygrotus impressopunctatus Pre 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 2 5 0 0 0
Hygrotus inaequalis Pre 0 17 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 9 144 0 0 0
Hygrotus marklini Pre 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 1 6 0 0 0
Hyphydrus ovatus Pre 0 3 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 31 0 0 0 0
Ilybius aenescens (larv) Pre 0 0 0 0 0 0 0 0 0 0 0 3 0 0 0 0 0 0 0 0 0 0
Ilybius fuliginosus (larv) Pre 0 0 0 0 3 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Ilybius sp. (larv) Pre 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 15 2 0 2
Laccobius bipunctatus Fin/Skr 0 0 0 0 0 0 0 0 0 0 0 0 3 0 0 0 0 0 0 2 0 0
Laccophilus minutus Pre 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 2 1 0 0 0
Limnebius crinifer Fin/Skr 2 0 0 0 0 1 0 4 0 0 0 0 0 2 7 0 0 2 0 0 0 0
Ochthebius minimus Fin/Skr 0 5 0 0 0 0 0 0 0 0 0 0 0 2 0 0 0 3 0 1 0 0
Porhydrus lineatus Pre 0 31 0 0 0 2 0 0 0 0 2 0 0 0 0 0 0 7 5 0 6 0
Rhantus frontalis Pre 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0
Rhantus suturalis Pre 0 0 0 1 0 1 0 0 0 0 2 1 0 1 0 1 0 1 1 0 0 0
Rhantus suturellus Pre 0 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 6 0 0 0 0
Trichoptera
Ek Ho Is Li Lu Mö Rö SjSH SjLH Sva Svä TA TB TC TD TE TF TG Tr Å1 Å2a Å2b Trichoptera
Limnephilus sp. Fra 0 0 0 0 2 1 0 0 0 0 15 0 1 0 0 0 0 0 0 5 0 0
Plectrocnemia conspersa Fil 2 7 0 0 0 0 0 0 0 0 0 0 0 0 0 0 9 0 0 0 0 0
Diptera Chaoboridae Pre 0 22 10 0 14 0 0 0 0 0 66 0 0 204 198 0 0 22 0 0 0 0 Chironomidae Fin 0 0 0 0 103 2 22 0 0 0 2 2 0 0 0 0 6 2 0 1 18 14 Culicidae Fin 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 Diptera Fin 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Sciomyzidae Pre 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Tipula sp. Fra 4 0 0 0 0 0 0 0 0 0 0 0 31 0 0 0 0 0 0 0 0 0 Isopoda
Asellus aquatiqus Fra/Fin 32 53 23 3795 11 3 13 216 50 3 91 26 52 227 158 63 0 676 1805 25 236 70
Amphipoda
Gammarus pulex Fra/Fin 548 0 0 0 0 6 0 28 64 0 0 0 134 237 156 0 412 0 0 0 0 0
Artantal per lokal 10 27 15 10 9 23 6 12 3 5 20 13 13 23 17 21 5 34 25 14 17 13
Tabell 8 Förklaringar till förkortningar
Fra Fragmenterare Ek Ekelunda Mö Möckelmossen Svä Svältbacken TE Tornrör E Å2a Åkerby källa 2a Fin Findetritusätare Ho Horsbrunn Rö Röhälla TA Tornrör A TF Tornrör F Å2b Åkerby källa 2b
Fil Filtrerare Is Ismantorp SjSH Sjukällorna Stora hålet TB Tornrör B TG Tornrör G Skr Skrapare Li Linnékällan SjLH Sjukällorna Lilla hålet TC Tornrör C Tr Triberga
3.5 Resultat av analys i Primer
En jämförelse av lokalernas likheter med varandra med avseende på evertebratfaunas sammansättning visade på ett antal grupperingar av källor (Figur 1). Grupperingarna var; Grupp 1 -Lund källa, Röhälla, Åkerby källa 1, 2a och 2b (Lund, Rö, Åk1, Åk2a och Åk2b) Grupp 2 - Åkerby källa 1 och Åkerby källa 2a och 2b (Åk1, Åk2a och Åk2b)
Grupp 3 - Horsbrunn, Svältbacken och Tornrör G (Hors, Svä och TG)
Grupp 4 - Stora hålet Sjukällorna, Tornrör C och Tornrör D (SHSj, TC och TD) Grupp 5 - Linnékällan och Triberga (Li och Tri)
Grupp 6 - Lund källa och Röhälla (Lund och Rö) Grupp 7 - Åkerby källa 2a och 2b (Åk2a och Åk2b) Grupp 8 - Svältbacken och Tornrör G (Svä och TG) Grupp 9 - Tornrör C och D (TC och TD)
Grupp 10 - Ismantorp och Tornrör E (Is och TE) Grupp 11 - Ekelunda och Tornrör B (Eke och TB)
Endast de lokaler som är lika varandra till minst 42 % har här ansetts vara grupper. Under denna gräns finns större grupperingar av källor, men de har i detta arbete bortsetts från då det inte går att utröna vilka de bakomliggande faktorerna till grupperingarna är. Gränsen 42 % är därmed inte baserat på någon typ av rekommendation utan bara utsatt att klargöra ungefär var gränsen går för resultatens förmåga att förklara grupperingarna övervägs av naturens komplexitet.
Lokalerna Möckelmossen (Mö), Tornrör A (TA), Åkerby Svartkällan (Sva), Tornrör F (TF) och Lilla Hålet Sjukällorna (LHSj) ligger ensamt fördelade mellan de andra grupperna.
Figur 1. Dendrogrammet visar grupperingen av lokaler. Lokaler i grupper under strecket är lika varandra med avseende på evertebratfaunan till minst 42 %.
Faktorer såsom inverkan av bete, beskuggning, andelen fin- och grovdetritus hade varken någon signifikant koppling till eller några mönster beträffande artantal och abundans. Det
fanns inte heller några signifikanta skillnader i artantal och abundans mellan de källor som grupperats efter närmiljö (alvar/skog/gräsmark).
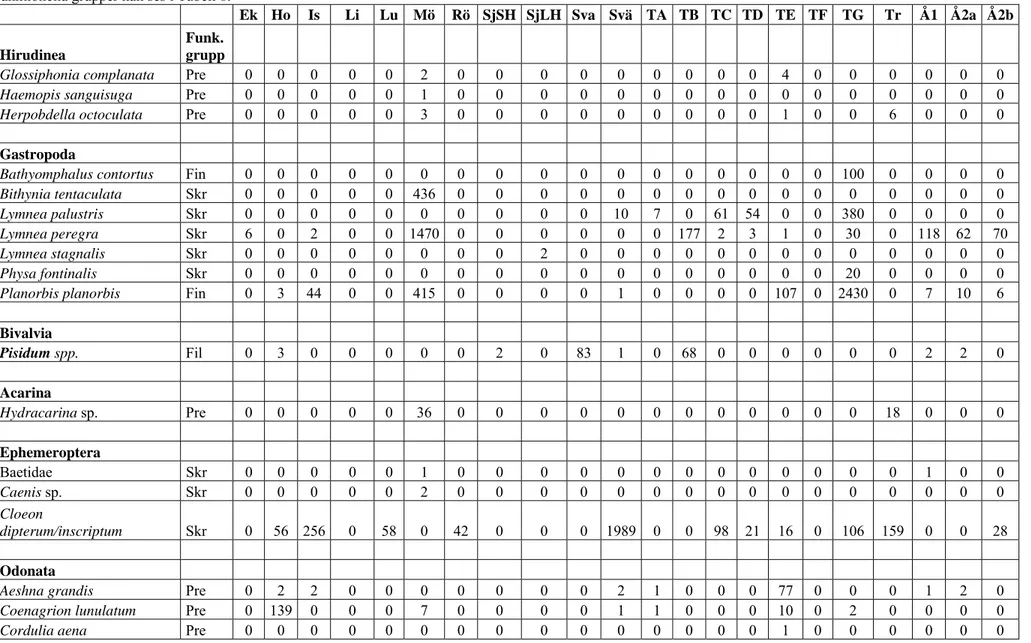
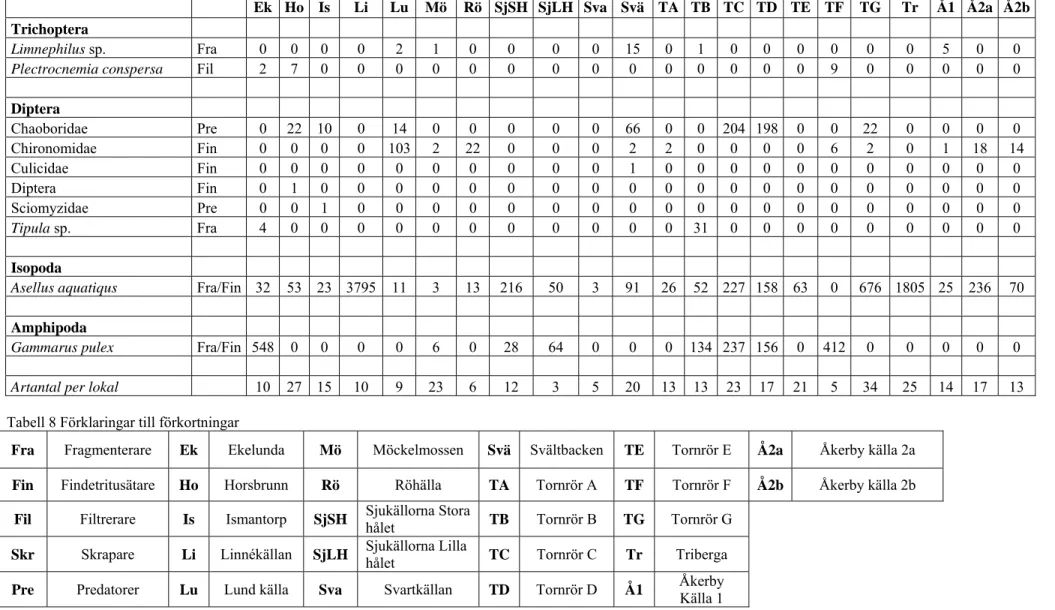
De faktorer som bäst förklarade variationen i det biologiska datamaterialet (Tabell 7) var omkrets, djup och beskuggning (corr. 0.439). De nämnda faktorerna var även tillsammans den bästa kombinationen av faktorer som förklarade variationen (corr. 0.439). Av de tre var det endast djupet som gav en signifikant skillnad då det studerades tillsammans med artantal och abundans (p = 0.023). Djupare källor hade således fler arter och individer av dessa än grundare källor (Figur 2 och 3).
Figur 2 och 3. Varje cirkel representerar en lokal och dess storlek är kopplat till abundans (antalet individer) och artantal dvs. ju större cirkel desto fler individer och arter finns inom lokalen. Graferna ger även en tvådimensionell uppfattning hur lika lokalerna är varandra där likheten mellan lokaler avspeglas i hur nära de ligger varandra. Således kan t.ex. i Figur 2 (t. v) ett mönster uppfattas där djupare källor som också har fler individer hamnar längre till vänster i grafen än de grundare källorna som ligger spridda till höger. För att se om ett mönster verkligen kan tänkas avspegla verkligheten görs även kompletterande signifikanstester, även om man ändå kan ta tillvara på tendenser till mönster som uppkommer i graferna så länge man inser att styrkan av dem är lägre om de saknar signifikans (se t.ex. Fig. 8-12). Dessa grafer visar att djupa källor har större abundans (Fig. 2, t. v) och ett större artantal (Fig. 3, t. h) än grunda källor, något som även bevisats med test av signifikans.
Även pH kunde signifikant förklara skillnader mellan lokalernas artantal och abundans (p = 0.012). I källor med högre pH var artantal och abundans högre än i de med lägre pH (Figur 4 och 5).
Figur 4 och 5. Källor med högre pH har större abundans (Fig. 4, t. v) och artantal (Fig. 5, t. h) än källor med lägre pH. Graferna tolkas på samma sätt som Figur 1 och 2.
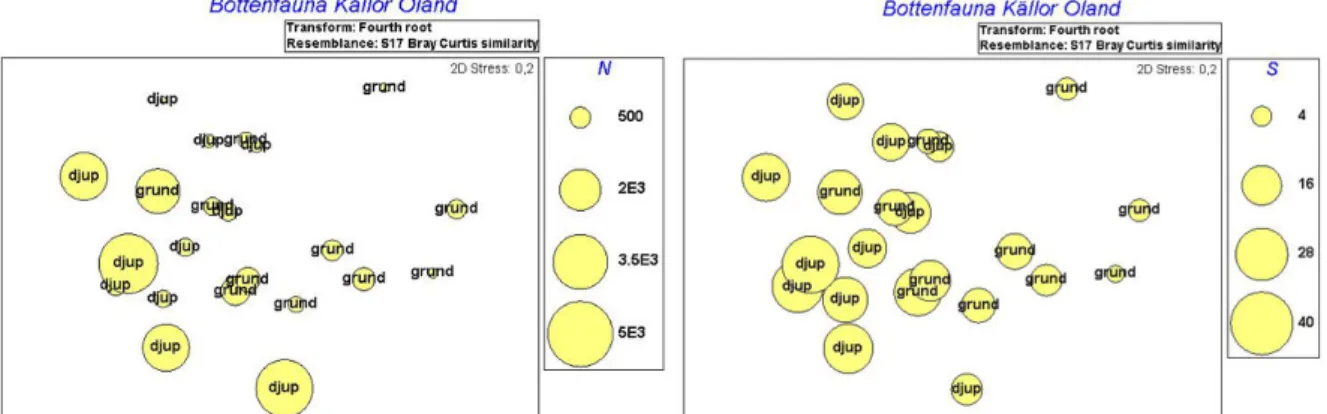
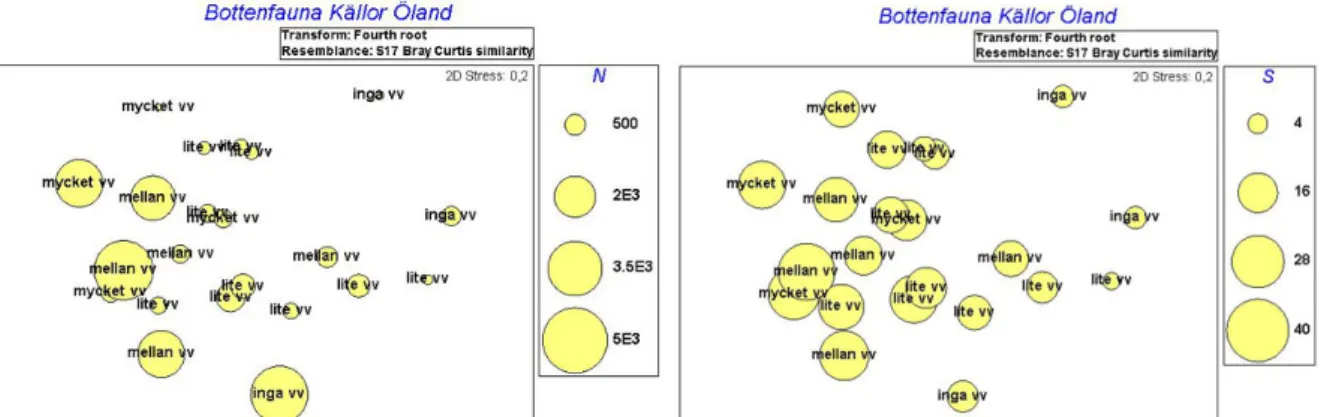
Figur 8 och 9. En tendens kan ses där abundansen (Fig. 8, t. v) och artantalet (Fig. 9, t. h) ökar med andelen vattenväxter. Mönstret avspeglar inte ett signifikant samband mellan abundans, artantal och andelen vattenväxter, i övrigt tolkas graferna så som beskrivits i Figur 1 och 2.
Figur 10 och 11. Graferna visar hur abundans (Fig. 10, t. v) och artantal (Fig. 11, t. h) kan tänkas öka med lägre konduktivitet. Mönstret avspeglar inte ett signifikant samband mellan abundans, artantal och konduktivitet, i övrigt tolkas graferna så som beskrivits i Figur 1 och 2.
Man kan även skönja ett mönster mellan storlek (omkrets) och artantal där större källor tycks ha fler arter (Figur 12), inget mönster fanns dock för abundans.
Figur 12. Grafen visar en tendens till fler arter i större källor (större omkrets). Mönstret avspeglar inte ett signifikant samband mellan artantal och källornas omkrets, i övrigt tolkas grafen så som beskrivits i Figur 1 och 2.
En korrelation mellan avståndsdata och data för lokalernas evertebratsamhälle visade att det fanns ett tydligt samband mellan avstånden mellan källorna och bottenfaunasamhället (p = 0.034). Vidare analys visade att källor skilda av mindre avstånd var mer lika varandra beträffande deras evertebratsamhällen än källor som skiljdes åt av större avstånd.
3.6 Funktionella grupper
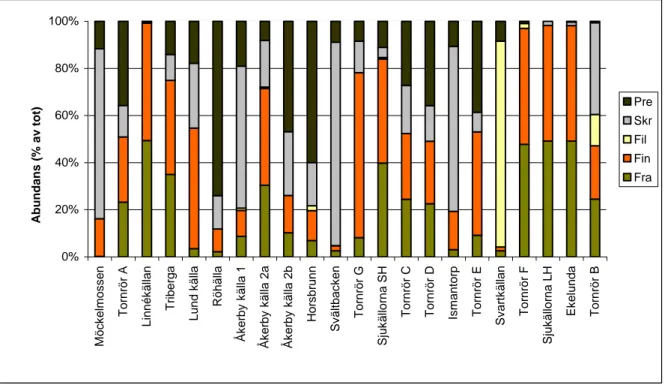
En jämförelse inom grupperingarna av källor som var lika varandra till minst 42 % vad gäller evertebratfaunan (Figur 1) och den procentuella fördelningen av de fem funktionella gruppernas abundans (Bilaga 1, Figur 1) och artantal (Bilaga 1, Figur 2) inom varje lokal, visade att de lokaler som hamnat i samma grupp hade en likartad artsammansättning mellan de funktionella grupperna inom varje lokal (ex Lund källa och Röhälla Bilaga 1, Figur 14). Filtrerare kunde dock saknas i vissa källor inom en gruppering av lokaler, men förekom de i mer än en källa per grupp fanns det likheter i andelen arter av skrapare mellan dessa källor. Däremot fanns inget sådant mönster för de funktionella gruppernas fördelade abundans inom vardera lokal med undantag för gruppen Tornrör C och Tornrör D (Bilaga 1, Figur 1). Källor i gräsmarks- och alvarmiljö hade en större andel individer av skrapare än källor i skogsmiljö, men andelen arter av samma grupp skiljde sig, liksom för andelen predatorarter, mindre åt mellan de tre olika miljöerna. Andelen individer av fragmenterare och findetritusätare dominerade ofta tillsammans i källor i skogsmiljö även om andelen individer av findetritusätare ofta var stor i källor i gräsmarks- och alvarmiljö. Andelen arter av fragmenterare, findetritusätare och predatorer skiljde sig inte mellan de tre olika miljöerna. Filtrerare förekom oftare och hade en större andel arter i källor i skogsmiljö. Andelen fragmenterare och findetritusätare ofta dominerade ofta med avseende på abundans i miljöer med träd runt om källan. Filtrerare utgjorde en lite större andel i dessa källor jämfört med lokaler med källor utan träd, där de dock förekom på fler lokaler. Fördelningen av artantalen mellan de funktionella grupperna skiljde sig inte mellan källor med eller utan träd. Större, medelstora och djupa källor hade en större andel skrapare abundansmässigt och en större andel predatorarter än mindre och grunda källor. Mindre och grunda källor hade en större andel individer av fragmenterare och findetritusätare, samt en högre andel arter av fragmenterare, än större och djupare källor. Källor med lägre pH hade en större andel arter av fragmenterare och findetritusätare tillsammans en högre andel av predatorer och filtrerare abundansmässigt. Andelen arter av predatorer och skrapare tycktes vara större i källor med högt pH.
4. Diskussion
Källor som grupperats av Primer hade stora likheter i fördelningen av de funktionella gruppernas arter men inte när det gällde dessa gruppers abundans med undantag av källorna Tornrör C och D (Avsnitt 3.5 Resultat av analys i Primer, Grupp 9). Och det var just källorna Tornrör C och D som fick mig att tro att det kanske var så att de kortare avstånden mellan källorna bidrog till att artsammansättningen av evertebrater i källorna blev så lika varandra men att de olika källmiljöerna sedan bestämde individantalen. Bara för att en art klarat av att migrera till nya områden betyder det inte att den kommer att klara sig lika bra i alla områden den tar sig till. Tornrör C och D låg inte bara närma varandra men deras miljöer var, till skillnad från de andra grupperingarna, i princip identiska vilket skulle kunna förklara varför man kunde se stora likheter i både artantal och abundans för dessa två källor men inte för de andra som endast var lika vad gäller artsammansättning. Det är alltså rimligt att tro, särskilt då det visade sig att källor skilda av kortare avstånd hade evertebratsamhällen som är mer lika varandra än källor skilda av större avstånd, att källornas artsammansättning av evertebrater påverkas av avstånden till andra källor men att individantalen av varje art sedan bestäms av
troligtvis förklarar varför de skiljde sig från andra källor. Möckelmossen var en av de största källorna medan Lilla Hålet Sjukällorna var en av de minsta. Tornrör A hade i sin tur en population av inplanterade guldfiskar som verkade trivas i källan året om och Svartkällan hade stora avlagringar av järnoxider och luktade stark av sulfid när sedimenten rördes om. Analysen av källor grupperade efter vilken typ av närmiljö de hade, visade att det inte fanns några signifikanta skillnader i artantal och antal individer mellan källor i skogsmiljö, alvarmiljö och de i gräsmark vilket betyder att de bentiska evertebratsamhällena i källorna som jag undersökt är mycket lika varandra. Även om alvarkällorna inte alltid hade träd runt sig hade de oftast, till skillnad från skogskällorna, en mer välutvecklad vegetation runtom och i källan vilket kan tänkas påverka evertebratsamhället. Makrofyter och mossor skapar heterogenitet i miljöer och ökar även den strukturella diversiteten (Ilmonen et. al. 2004). Effekten av detta blir i sin tur en ökning av den biologiska mångfalden (Lindegaard et. al. 1998). Jag tror därför att alvarkällors status som potentiell refug för bl.a. växter i en annars torr och näringsfattig miljö till viss del kan förklara varför de inte skiljer sig från källor i skogsmiljö beträffande artsammansättning av evertebrater. Källor som låg i gräsmarker hade dels den översvämmade gräsmarken, men också ibland träd eller buskar i närheten av källan, samt en mindre andel vattenväxter som utgjorde vegetationen runt och i källan. Deras likhet med källor i alvar- och skogsmiljö kan bero på att de både är öppna med tämligen mycket vegetation (liksom källor i alvarmiljö) men även har inslag av träd och buskar (liksom källor i skogsmiljö).
Jag undersökte även fler faktorer som verkade kunna förklara skillnader i artantal och abundans, såsom djup, pH, förekomsten av vattenväxter, konduktivitet och källornas storlek. Ett mönster kunde urskiljas där en mer dominant vegetation av vattenväxter ledde till högre artantal och abundans vilket kan förväntas då en större andel växter leder till ökad biologisk mångfald. Att artantal och abundans ökar med djup och möjligen även med källornas omkrets är inte alltför förvånande då både djup och omkrets bidrar till källans storlek, som enligt den öbiogeografiska teorin utgör en av faktorerna som kan ge en miljö möjlighet att hysa fler potentiella arter då den ökar (Frick et. al. 2008). Vattenlevande organismer har olika preferenser för pH i deras livsmiljö och kanske finns det fler arter och högre abundans i de källor som har högre pH då de organismer som lever i källor på Öland möjligtvis trivs bättre i och kanske även är mer anpassade till de kalkrikare miljöer som Öland oftast representerar. Det fanns dock inga av mina resultat som visade att det var endast alvarkällor som överlag var djupare, hade större omkrets eller hade högre pH. Det fanns ett mönster som visade ett större artantal och en högre abundans i källor med lägre konduktivitet vilket snarare beror på tillgången till syre och inte konduktiviteten i sig. Distributionen av många typer av organismer, däribland evertebrater, är starkt beroende av tillgången till syre som kan ses som en begränsande faktor (Van der Valk 2007). En ökning i primärproduktionen kan senare leda till syrebrist då det producerade materialet ska brytas ned. Vid nedbrytningen av det organiska materialet ökar mängden lösta joner i vattnet vilket i sig leder till en ökning av konduktivitet. Således kan en hög konduktivitet vittna om en ökad nedbrytning och minskad syrehalt som är en styrande faktor för distributionen av olika typer organismer.
Fragmenterare och findetritusätare dominerade abundansmässigt i mindre och grundare källor där predatorer ibland saknades eller fanns i lägre antal. De större och djupare källorna hade fler predatorarter som kunde hålla efter sina potentiella bytesdjur. Källor i alvarmiljö hade tex en jämnare fördelning i abundans av de funktionella grupperna än källor i skogs- och gräsmarksmiljö, men i de fall som andelen individer av predatorer är mindre ökar de andra grupperna, främst findetritusätarna och fragmenterarna. Den mindre andelen predatorer i de
mindre och grundare källorna kan därför tänkas vara orsaken till att fragmenterare och findetritusätare ofta dominerade. Förklaringen till skillnaderna i andelen predatorarter mellan källorna kan vara att många predatorer föredrar miljöer med en större permanens i vattenförekomst då många, men inte alla (Boix et. al. 2006), är känsliga för uttorkning (Wellborn et. al. 1996). Detta skulle kunna vara förklaringen till att predatorer ofta saknades eller fanns i ett mindre antal/artantal i mindre källor, men eftersom källor sällan eller aldrig torkar ut finns det troligtvis en annan förklaring till att de föredrar de större källorna. Det kan vara så att fördelningen av predatorarter inte beror på källornas djup utan är en effekt av källornas storlek förklarat av den öbiogeografiska teorin. Källorna i gräsmarksmiljö var som sagt ofta öppna miljöer med översvämmade ytor med gräs och halvgräs. På denna vegetation satt ofta påväxtalger, troligtvis till följd av att mycket solljus tilläts nå källmiljön på grund av öppenheten, och jag blev inte förvånad när andelen individer av skrapare var mycket högt på dessa lokaler troligtvis på grund av fördelen av de stora födoresurserna. Även alvarkällor hade en stor andel individer av skrapare, här troligtvis också som en effekt av födoresurser kopplade till andelen solljus som når källmiljön. Det fanns vissa trender för de funktionella grupperna med avseende på hur de fördelade sig efter pH men dessa förklaras nog bäst av de ingående arternas individuella krav på vattenkemi, något som inte tas upp i detta arbete men som kanske skulle kunna vara intressant att undersöka. En av de minsta funktionella grupperna var i de undersökta källorna filtrerarna som endast representerades av två arter (Ärtmussla Pisidum spp. och en nattslända Plectrocnemia conspersa) och just därför kan man kanske bara säga att det antal och särskilt artantal filtrerare jag funnit visar på att denna funktionella grupp kanske inte trivs i de källor jag undersökt. En undersökning av bl.a. tillgången på planktoniska alger skulle möjligtvis kunna berätta mer om varför denna grupp förekommer så sparsamt.
5. Sammanfattning
Då det inte finns några tidigare studier av den bentiska evertebratfaunan i källor på Öland ger detta arbete en första inblick i denna miljö. Det finns fortfarande många frågetecken om vad som bestämmer utformningen av källornas evertebratsamhällen, samt fördelningen av arter och abundans av de funktionella grupperna, även om jag har tagit upp några tänkbara teorier baserat på de resultat jag erhållit. Arbetet kan behöva kompletteras särskilt med en uppföljning av evertebratfaunans artsammansättning och abundanser under våren eftersom detta arbete utfördes sent på året.
6. Tack
Tack till Jan Herrmann, Börje Ekstam och Geoffrey Lemdahl för god handledning, kritik och uppmuntran. Staffan Arvegård, Roland Bengtsson, Anders Eklöv, Thorsten Jansson, Anders Johansson, Tommy Knutsson och Eje Rosén för tips och hjälp att lokalisera källor. Roland Engkvist och Patrik Dinnetz för förslag till och handledning vid statistiska analyser. Joja Geijer för hjälp med och handledning vid nyckling av skalbaggar. Karolina Leberfinger för hjälp med nyckling av nattsländelarver. Berit Hilmersson, Anna Hilmersson, Torsten Hilmersson och Sofie Jansson för assistans i fält och kärleksfullt stöd. Lokalbefolkningen på Öland för det trevliga bemötandet och given hjälp då denna kunde ges.
7. Referenser
Boix D, Sala J, Gascon S, Brucet S, 2006, Predation in a temporary pond with special attention to the trophic role of Triops cancriformis (Crustacea: Branchiopoda: Notostraca), Hydrobiologia vol 571 pp 341–353 Bråvander L-G, Wiklander G, 2006, Livet i Källorna – I: Håkan S (red); Källor i Sverige pp. 55-76, AB Svensk Byggtjänst, Sundbyberg, Sverige
Edington J M, Hildrew A G, 1995, A revised key to the Caseless caddis larvae of theBrittish Isles with note on thier ecology, Freshwater biological association, scientific pulication No. 53
Enckell P H, 1980, Kräftdjur, Bokförlaget Signum, Lund, Sverige
Engelhardt Wolfgang, 1976, The young scientists looks at at Pond Life, Burke publishing Company, Storbritannien
Eriksson A, Wiklander G, 2006, Skydd och vård av Källor – I: Håkan S (red); Källor i Sverige pp. 167-174, AB Svensk Byggtjänst, Sundbyberg, Sverige
Frick W F, Hayes J P, Heady P A, 2008, Island biogeography of bats in BajaCalifornia, Mexico: patterns of bat speciesrichness in a near-shore archipelago, Journal of Biogeography vol 35 pp 353-364
Habdija I, 2001, Tufa deposition and periphyton overgrowth as factors affecting the ciliate community on travertine barriers in different current velocity conditions, Hydrobiologia vol 457 pp 87–96
Hoffsten P-O, 2006, Livet i Källorna – I: Håkan S (red); Källor i Sverige pp. 55-76, AB Svensk Byggtjänst, Sundbyberg, Sverige
Hubendick B, 1949, Våra snäckor - Snäckor i sött och bräckt vatten, Bonniers, Stockholm, Sverige
Hult A, 2006, Källvattnets bildning och egenskaper – I: Håkan S (red); Källor i Sverige pp. 13-28, AB Svensk Byggtjänst, Sundbyberg, Sverige
Ilmonen J, Paasivirta L, 2004, Benthic macrocrustaceans and insect assemblages in relation to spring habitat characteristics: patterns in abundance and diversity, Hydrobiologia vol 533 pp 99-113
Knutsson G, 2006a, Källvattnets bildning och egenskaper – I: Håkan S (red); Källor i Sverige pp. 13-28, AB Svensk Byggtjänst, Sundbyberg, Sverige
Knutsson G, 2006b, Olika typer av källor – I: Håkan S (red); Källor i Sverige, pp. 29-44 AB Svensk Byggtjänst, Sundbyberg, Sverige
Knutsson G, 2006c, Förändringar i och kring källor – I: Håkan S (red); Källor i Sverige pp. 45-55, AB Svensk Byggtjänst, Sundbyberg, Sverige
Landin B O, 1970, Insekter, A&W, Stockholm, Sverige
Lindegaard C, Brodersen K P, Wiberg-Larsen P, Skriver J, 1998, Multivariate analyses of macrofaunal communities in Danish springs and springbrooks – I: Botosaneanu L (editor); Studies in crenobiology pp. 201-219, Backhuys Publishers Leiden, Nederländerna
Løjtnant B et al, 1990, Danske Vandplanter – Vejledning i bestemmelse af planter i søer og vandløb, Miljønyt Nr. 2, Miljøministeriet och Danmarks Miljøundersøgelser, Danmark
Mandahl-Barth G, 1970, Vad jag finner i sjö och å, A&W, Stockholm, Sverige Mandahl-Barth G, 2000, Småkryp i Sötvatten, Fältbiologernas förlag, Sverige
Meier-Brook C, Glöer P, 1998, Süsswassermollusken – Ein bestimmungsschüssel für die Bundesrepublik Deutschland, Deutscher Jugendbund für Naturbeobachtung DJN, Hamburg, Tyskland
Mossberg B, Stenberg L, 2003, Den nya nordiska floran, Wahlström &Widstrand, Tangen, Norge Naturvårdsverket, 2001, Undersökningstyp - Lokalbeskrivning, Handbok för miljöövervakning
Nilsson A, 1996, Aquatic Insects of North Europé vol. 1 – A taxonomic handbook, Apollo Books, Stenstrup, Danmark
Nilsson A, 1997, Aquatic Insects of North Europé vol. 2 – A taxonomic handbook, Apollo Books, Stenstrup, Danmark
Sahlén G, 1996, Sveriges Trollsländor – En bestämningsbok för trollsländor i Sverige och övriga Norden, Fältbiologernas förlag, Stockholm, Sverige
Savage A A, 1989, Adults of the British aquatic hemiptera heteroptera; a key with ecological notes, Freshwater Biological Association, Scientific Publication No. 50
Svensson B, 1980, Akvatiska Dipter-larver i Sverige – Bestämningsnyckel för familjer; Tipulidae, Cylindrotomidae & Limoniidae, Stencil
Svensson B S, 1986, Sveriges dagsländor (Ephemeroptera) - Bestämning av larver, Ent. Tidskr. 107:91-106, Umeå, Sverige
Van der Valk A G, 2007, The biology of freshwater wetlands, Oxford University Press Inc, New York, USA Wallace B, Wallace I D, Philipson G N, Keys to the Case-bearing caddis larvae of Britain and Ireland, 2003, Freshwater biological association, scientific pulication No. 61
Wellborn, G A., Skelly, D K, Werner E E, 1996. Mechanisms creating community structure across a freshwater habitat gradient. Annual Revision of Ecological Systematics 27: 337-63.
Wikipedia, 2007, http://sv.wikipedia.org/wiki/Limonit,15.49, 2008-02-05
Bilaga 1. Vattenkemi, bottensubstrat, markanvändning, närmiljö och vegetation Lokalbeskrivningar
Ekelunda
Miljön runt källan består av Alvar med tidvis täta bestånd av en och enstaka träd av björk, tall olvon och slån. Marken används för bete av kor. Källan i sig är inte stor eller särskilt djup och är till hälften belägen under en buske. Från källuppflödet mynnar en mindre källbäck som löper ut i ett anslutande vattendrag. Markvegetationen runt om källan består till största delen av gräsmark.
Horsbrunn
Horsbrunn, eller Horskällan som den även kallas, är en större källa belägen i lövskog mestadels bestående av björk och hassel men även sälj och al. Vegetationen runt om källan präglas av halvgräs och småbuskar av bl.a. sälj. I källan finns dels undervattensvegetation i form av kransalger samt flytbladsväxter. Ett träd som trillat ligger till hälften nedsänkt i källans ena parti och en stor del av källans organiska material består av löv i olika nedbrytningsstadier.
Ismantorps borg
Själv källan är belägen en bit utanför borgruinen. Gräsmarken med sina spridda enbuskar samt bestånd och skogspartier av framförallt ek men även gran används idag som bete för får. Källans kanter och i viss mån mitt präglas av övervattensvegetation och några flytbladsväxter. Linnékällan
Denna djupa källa saknar vattenvegetation med undantag av den andmat man finner på dess yta. Kanterna på källan består av raka kalkstensväggar utan några större ansamlingar av organiskt material. Marken runt källan domineras av gräs och används för bete av kor. Inga träd eller buskar finns i källans närmiljö, denna präglas istället av betesmarker för boskap samt åkrar.
Lund källa
Lund källa är placerad på Karums alvar. Källans närmiljö består av alvar- och hällmark med fårbete med inslag av skog och enskilda bestånd av träd såsom oxel, björk och tall samt buskar av t.ex. slån, en, hassel, nypon och björnbär. Några träd står runt kanten på källan vars omgivning domineras friskare gräsmark med inslag av örter. En del flytbladsväxter och undervattensväxter finns i källan och ute i den fria vattenmassan syns stora sjok av grönalger som även verkar finnas nedsjunkna till botten.
Möckelmossen
Källan är belägen en bit sydöst om den iordningställda parkeringen till den stora mossen. Omgivningen präglas av alvarmark med enstaka enar och frisk gräsmark samt häll runtom källan. Vattnet utnyttjas av betande kor som till viss del trampat upp gräsmarken runt källan och som lämnat efter sig avföring i nära anslutning till eller i källans vatten. Vegetationen i
källan består av flytbladsväxter, kransalger och långskottsväxter och bottensubstratet består av ett tämligen tjockt lager dy ovanpå hällen som annars utgör källans botten.
Röhälla
Källan i Röhälla ligger på gräsmark som betas av kor. I anslutning till källan står ett fåtal träd men en längre bit bort finns skog av bl.a. tall. Påverkan av korna är tydlig då marken är påtagligt upptrampad och korna kan ses göra sina behov i eller bredvid källan då de besöker den för att dricka. Källan är mycket dyig och vegetationen domineras av gräs och en del andmat samt ett fåtal örter.
Sjukällorna
Sjukällorna består av två, kanske tre separata uppflöden av källvatten, två som bildar var sitt källkar och en som verkar bryta fram på bred front och skapa ett blötare område inom lokalen. Miljön runt om består av gräs- och halvgräs med anslutande skog av bl.a. björk med inslag av enbestånd. Marken betas av får och kor men dessa verkar dricka från vattendraget som bildas då de 3 källflödena sammanfaller till ett större vattendrag. Vegetationen i källorna är mycket sparsam och består till största delen av grönalger. Det ena källflödet sjuder på grund av vattenströmningen medan de båda andra verkar flyta fram långsammare. Således finns det mindre organiskt material ansamlat i den sjudande källan, medan det andra källkaret har ansamlingar av löv.
Svartkällan
Källan är belägen i ett mindre skogsbestånd av bl.a. hassel och asp. Järnutfällningar kan ses på löv och annat organiskt material som ansamlats i källan. Vid omrörning av vattnet känns en lukt av ruttna ägg och vattnet färgas svart av bottenmaterialets sulfid. Runt om källan finns en del buskar av bl.a. björnbär. Det ursprungliga källflödet har provborrats vid prospekteringar av vattentäkt och består därför av en brunn med lock som ibland svämmar över vid mycket regn. Marken runt beståndet av träd används som åker.
Svältbacken
Ett eller flera källflöden bildar tre olikstora källkar i den friska marken som domineras av halvgräs och mossa. Marken används som bete till hästar som dricker källans vatten. Omgivningarna är dock ej avsevärt upptrampade sin friskhet till trots men en mindre mängd spillning kan ses i källans vatten. I källans närmiljö finns en del skog sammansatt av ek och björk med inslag av en. Kalkutfällningar och påväxtalger kan ses på vegetationen i och runt om källan.
Tornrör A-F
Tornrör A – Denna källa omges av en miljö präglad av skog, alvarmark och gräsmark. Runtom källan är marken täckt av gräs och halvgräs med inslag av örter något som sedan övergår i en blandning av skog och alvar med tidvis täta bestånd av en och hassel. Det dominerande trädslaget är björk. Själva källan rinner upp i en punkt varifrån vattnet sedan
Tornrör B – Den så kallade Tornrörskällan bryter fram under ett bestånd hassel och har stenlagts. Nedanför stenläggningen bildar vattnet ett mindre källkar med dyig botten ovanpå kalkstensgrunden. Vegetationen domineras av halvgräs och gräs med en del påväxtalger. Runt om källan är marken frisk till sank med en- och slånbuskar på de torrare partierna. Löv från hasselbeståndet kan ses flyta ner i källkaret men några ansamlingar av löv kan inte ses. Den friska marken är på sina ställen upptrampad av boskap och avföring kan ses i vattnets närhet. Tornrör CD – Lokalen CD innefattar två skilda källor som ligger med ca 3 m avstånd från varandra där källa C ligger något söder om källa D. Den miljön runt om och i källorna är mycket lika, därför kommer den beskrivande texten att omfatta dem båda. Marken runt om källorna domineras nästan helt uteslutande av halvgräs och är på sina ställen upptrampade av boskap. Botten i källornas yttre kanter är en ibland dyig kalkstenshäll med grunt vatten som när det blir djupare mot mitten bryts upp i större bitar av kalksten som kan ses där vegetationen saknas. I källorna finns en del vattenvegetation runt kanterna och en bit ut mot mitten med inslag av örter och utfällningar av kalk kan ses på växterna.
Tornrör E – Runt kanten på halva källan finns skog av björk och hassel med inslag av en. Runt resten av källan finns frisk mark dominerad av gräs, halvgräs och örter som är upptrampad av boskap som även lämnat avföring i vattnet. En bit bort från källan öppnar sig alvaret med spridda enbuskar. Källans botten är dyig med ansamlingar av löv ovanpå den flata kalkstensberggrunden som verkar ha sprängts upp vid källuppfödet för att göra källan djupare. Sprängmassor ligger vid sidan av källan vars djup är etapplikt och mycket djupt i närheten av det misstänkta källuppflödet. I den djupare delen finns en del flytbladsväxter, annars är det rosettväxter, övervattensväxter och halvgräs som dominerar vegetationen i källan.
Tornrör F – Källan ligger i en dunge av hassel- och enbuskar samt björkar vars barr och löv kan ses ansamlade i källflödet. Vegetation i och runt källan saknas för övrigt kanske med undantag för lite grönalger på stenarna i vattnet. Bottnen består av en nästan plan kalkstenshäll med ett tunt sandlager med inslag av dy. Marken runt källan är upptrampad av boskap och avföring från kor finns runt om och ibland i källan.
Tornrör G
Denna källa ligger inte inom lokalen Tornrör men hittades av en slump då lokalen Tornrör skulle besökas. Eftersom denna lokal ligger runt 1 km från det riktiga Tornrör och annars saknade namn på kartan fick den tillhöra Tornrör namnmässigt.
Miljön runt källan består av tidvis frisk mark som domineras av halvgräs och gräs på vilket man kan se kalkutfällningar. Spridda ansamlingar av träd och buskar med björk, ek, hassel och en kan ses i området som i olika riktningar övergår i alvar, bete för kor, åker och skog. Källan har fördjupats via grävning, troligtvis för vattning av boskap, något som kan ses på de två högar som ligger i anslutning till vattnet. I källan finns en del flytbladsväxter och kransalger, längs kanten växter gräs, halvgräs och lite örter. Bottnen är något dyig och sluttar tämligen starkt mot mitten.
Triberga borg
Källan är placerad innanför fornborgens murar i en rektangulär nedsänkning i marken med raka kanter av kalksten som har en del gräs i sina sprickor. En liten sluttning finns i ena sidan av källan där borgens dåtida invånare kunde hämta vatten. Vegetationen i källan domineras av kransalger som bildar en matta på dess botten. Marken ovanför källan domineras av gräs inuti
borgen med alvarmark utanför, träd saknas. Källan har inga större ansamlingar av organiskt material och bottensubstratet består av större bitar av kalksten.
Åkerby källor
Den källa som fått namnet ”Åkerby källa” är omgiven av frisk och tidvis sank mark som domineras av halvgräs och mossa. Den friska marken som betas av hästar är kantad av blandskog av bl.a. björk, en och gran. Påväxtalger och kalkutfällningar kan ses på vegetationen i och vid källan. Den ursprungliga källan har borrats vid en vattenprospektering och stenlagts, men verkar ändå vara centrum för uppflödet av vatten även om man kan se vatten sippra upp i marken runtom brunnen. I närheten av källan hittades skal av kräftor som tros härstamma från en kräftodling nedströms. (Åkerby källa 2a och 2b)
Ytterligare en källa hittades runt 100 m från Åkerby källa. Denna källas omgivande miljö skiljde sig inte nämnvärt frånsett det faktum att den låg närmare den omgivande skogens kant. Även här kunde man se kalkutfällningar och påväxtalger på vegetationen vid källan. Marken runt källan är frisk men inte lika sank som den runt Åkerby källa och domineras nästan enbart av halvgräs med ett mindre inslag av mossa. (Åkerby källa 1)
Vattenkemi, bottensubstrat, markanvändning, närmiljö och vegetation
Tabell 1. Vattenegenskaper och källstorlek. Då det ej gått att mäta djup fullt ut har en uppskattning gjorts som borde avspegla de egentliga djupen.
pH kond (µs/cm)
vatten-hastighet vattenfärg grumlighet
omkrets (m) medeldjup (m) maxdjup (m)
Ekelunda 7.24 486 1 klart klart 19 0.07 0.1 Horsbrunn 7.76 326 0 klart klart 100 0.5 1.4 Ismantorp 7.52 530 0 klart klart 18 0.45 0.6 Linnékällan 7.5 725 0 färgat klart 10 2 2 Lund källa 7.38 568 0 färgat grumligt 57 1 1.5 Möckelmossen 8.11 236 0 klart klart 70 0.4 1 Röhälla 6.46 300 0 klart klart 49 0.35 0.5 Sjukällorna SH 7.2 487 1 klart klart 8 0.4 0.45 Sjukällorna LH 7.2 487 2 klart klart 4 0.18 0.3 Svartkällan 6.98 798 1 färgat klart 8 0.3 0.6 Svältbacken 7.82 445 0 klart klart 39 0.25 0.4 Tornrör A 7.2 526 1 färgat klart 38 0.7 2 Tornrör B 7.27 534 1 färgat klart 28 0.07 0.1 Tornrör CN 7.71 492 0 klart klart 16 0.25 0.7 Tornrör DS 7.61 545 0 klart klart 19 0.3 0.7 Tornrör E 7.08 514 0 klart klart 41 0.5 0.75 Tornrör F 7.07 520 1 klart klart 28 0.1 0.16 Tornrör G 8.83 182 0 klart klart 27 0.5 0.75 Triberga 8.07 276 0 klart klart 18 0.5 1 Åkerby källa 1 7.37 632 1 klart klart 18 0.1 0.75 Åkerby källa 2A 7.12 652 1 klart klart 14 0.35 0.4 Åkerby källa 2B 7.12 652 1 klart klart 33 0.2 0.2
Tabell 2. Bottensubstrat, detritus och död ved. Hur substratens dominans varierar i förhållande till varandra är baserat på en tre-gradig skala efter vilken en uppskattning av förekomsten av de olika substratens gjorts.
fin-sediment sand grus fin sten grov sten fina block grova block häll fin detritus grov detritus fin död ved grov död ved Ekelunda 1 0 2 1 1 2 0 0 1 2 0 0 Horsbrunn 0 0 1 0 0 0 0 3 2 3 1 2 Ismantorp 0 0 1 2 3 0 0 0 1 3 0 0 Linnékällan 0 0 0 0 0 0 0 0 2 2 0 0 Lund källa 0 0 1 1 2 2 0 0 3 1 0 0 Möckelmossen 1 0 1 1 1 1 0 2 2 3 0 0 Röhälla 0 3 0 0 0 0 0 0 3 1 0 0 Sjukällorna SH 1 2 3 0 0 0 0 1 2 3 0 0 Sjukällorna LH 1 2 3 0 0 0 0 1 1 1 0 0 Svartkällan 0 0 2 1 1 2 1 0 1 2 0 2 Svältbacken 0 0 0 0 3 2 0 0 1 2 0 0 Tornrör A 0 0 2 2 2 1 0 0 2 2 0 0 Tornrör B 0 0 1 1 1 2 0 3 3 2 0 0 Tornrör CN 0 0 0 1 0 3 0 2 1 1 0 0 Tornrör DS 0 0 2 0 0 1 0 3 2 1 0 0 Tornrör E 0 1 1 0 0 1 2 3 1 3 1 0 Tornrör F 0 0 2 2 1 1 2 1 1 3 0 0 Tornrör G 0 0 2 2 1 1 0 1 1 2 0 0 Triberga 1 0 1 0 0 1 2 2 3 1 0 0 Åkerby källa 1 0 0 0 0 0 0 0 0 3 1 0 0 Åkerby källa 2A 0 0 0 0 1 2 0 3 2 2 0 0 Åkerby källa 2B 0 0 0 0 0 1 0 3 2 2 0 0
Tabell 3. Buskar och träd samt, markanvändning, marktyp och grad av beskuggning. Förklaring till förkortningar kan ses i tabell 4.
mark- användning marktyper beskugg -ning buskar och träd dom. arter buskar och träd sub. dom arter Ekelunda bete kor alvar, blandskog 1 B, En T, Sl ,Ol Horsbrunn skog blandskog 3 B, H Sä, A Ismantorp bete får äng, lövskog 1 Ek, En B, H Linnékällan Bete kor äng 0
Lund källa bete får alvar, blandskog 2 Sl, B, Ox, En, H T, N, Bb Möckelmossen bete kor alvar 0 En
Röhälla bete kor äng, blandskog 0 T Ox, B Sjukällorna SH bete kor och får alvar, blandskog 2 B, En G, T Sjukällorna LH bete kor och får alvar, blandskog 2 B, En G, T Svartkällan skog lövskog, åker 3 As, H Bb Svältbacken bete häst äng, lövskog, våtmark 0 B, Ek En Tornrör A bete kor alvar, lövskog 0 En, H Ö Tornrör B bete kor alvar, lövskog 1 B, En Sl, H Tornrör CN bete kor alvar 0 En H Tornrör DS bete kor alvar 0 En H Tornrör E bete kor alvar, lövskog 2 B, H En Tornrör F bete kor alvar, lövskog 3 B, H En Tornrör G bete kor alvar, våtmark 0 B, En Ek, H, Sl Triberga fornborg alvar 0
Åkerby källa 1 bete häst och kor blandskog, våtmark 0 G, B, En Ek, Sä, H Åkerby källa
2A bete häst och kor blandskog, våtmark 0 B, En G Åkerby källa
2B bete häst och kor blandskog, våtmark 0 B, En G Tabell 4. Förklaring till förkortningar i Tabell 3.
A-al (Alnussp.) N-nypon (Rosa sp.) As-asp (Populus tremula) Ol-olvon (Viburnum opulus) Bb-björnbär (Rubus sp.) Ox-oxel (Sorbus sp.) B-björk (Betula sp.) Sl-slån (Prunus spinosa) Ek-ek (Quercus robur) Sä-sälg (Salix caprea) En-en (Juniperus communis) T-tall (Pinus sylvestris)
G-gran (Picea abies) Ö-ölandstok (Dasiphora fruticosa) H-hassel (Corylus avellana)
Tabell 5. Vattenvegetation, mossor och alger övervatten-växter flytblads-växter långskotts-växter
rosett-växter mossor alger
Ekelunda 0 0 0 2 1 0 Horsbrunn 2 2 0 0 2 1 Ismantorp 2 1 0 0 0 0 Linnékällan 0 3 0 0 0 0 Lund källa 1 0 0 0 1 0 Möckelmossen 3 2 1 2 1 1 Röhälla 2 2 0 1 0 1 Sjukällorna SH 1 0 0 0 1 1 Sjukällorna LH 1 0 0 0 1 3 Svartkällan 3 0 1 0 0 0 Svältbacken 3 0 0 0 2 1 Tornrör A 1 2 2 1 1 2 Tornrör B 1 0 1 1 1 3 Tornrör CN 2 0 2 1 1 1 Tornrör DS 2 0 2 1 1 1 Tornrör E 2 2 1 1 1 0 Tornrör F 0 0 0 0 0 0 Tornrör G 1 2 1 1 0 0 Triberga 2 0 0 1 3 0 Åkerby källa 1 2 0 0 0 2 3 Åkerby källa 2A 3 0 1 1 2 1 Åkerby källa 2B 3 0 1 1 2 1
Tabell 6. Förekomst/Icke förekomst av de dominerande växt- och mossarterna i och runt källan. Förklaringar till lokalförkortningar finns i Tabell 8. Ek Ho Is Li Lu Mö Rö SjSH SjLH Sva Svä TA TB TC TD TE TF TG Tr Å1 Å2a Å2b Blysmus compressa x Calliergon cordifolium x x x x x Carex sp. x x Chara sp. x x x x x x x x x x Chlorophyta x x Chrysoplenium alternifolium x Cyperaceae x x Drepanocladus fluitans x Equisetum fluviatile x x Equisetum pratense x x Fontinalis antipyretica x x x Fontinalis dalecarlica x Juncus articulatus x x x x x x x Juncus sp. x Mentha arvensis x x x x Menyanthes trifoliata x x x x x Lemna sp. x x x Pleurozium schreberi x x x Potamogeton natans x x x x Ranunculus aquatilis x x Rhytidiadelphus sqarrosus x x Schoenoplectus lacustris x Scorpidium scorpioides x x x x Sesleria caerulea x x x x x x x x x