Svensk Botanisk
Tidskrift
Utgiven av
Svenska Botaniska Föreningen
Redigerad av
STEN AHLNER
BAND 51
1957
HÄFTE 4SVENSKÅ BOTANISKA FÖRENINGENS
styrelse och redaktionskommitté år 1957.
Styrelse:
E. MELIN, ordförande; R. FLORIN, v. ordförande; G. HARLING, sek reterare; S. AHLNER, redaktör och ansvarig utgivare av tidskriften; C.-A. TORÉN, skattmästare; I. HOLMGREN, E. HULTÉN, T. LAGER BERG, C. MALMSTRÖM, J. A. NANNFELDT, M. G. STÅLFELT,
H. WEIMARCK. Redaktionskommitté:
G. E. DU RIETZ, E. HULTÉN, T. LAGERBERG, C. MALMSTRÖM, J. A. NANNFELDT, M. G. STÅLFELT.
SVENSK BOTANISK TIDSKRIFT utkommer med fyra häften årligen. Prenumerationsavgiften (för personer, som ej tillhöra Svenska Botaniska
Föreningen) är 25 kronor. Svenska och utländska bokhandlare kunna direkt hos föreningen erhålla tidskriften till samma pris.
Medlemsavgiften, för vilken även tidskriften erhålles, är 20 kronor för medlemmar, bosatta i Sverige, Danmark, Finland, Island och Norge, och kan insättas på föreningens postgirokonto 2986 (giroblankett åtföljer häfte nr 1 för året) eller översändas på annat sätt. Har så ej skett före ut givandet av häfte nr 2, utsändes detta mot postförskott, varvid porto debiteras. Medlemmar erhålla i mån av tillgång tidigare årgångar av tidskriften till ett pris av 16 kronor per årgång.
Generalregister över de första 40 årgångarna finnas nu tillgängliga.
SVENSK BOTANISK TIDSKRIFT, edited by Svenska Botaniska Föreningen (The Swedish Botanical Society), is issued quarterly.
An annual fee of 25 Sw. Kr., which includes the journal, applies to mem bers outside Sweden, Denmark, Finland, Iceland and Norway. The jour nal is available to booksellers for the same amount. Back volumes are available to members at 16 Sw. Kr. according to supply.
A general index, in two parts, to Volumes 1—40 is now available.
DE MÖRKRÖDA MITTBLOMMORNA IIOS
VILD MOROT.
AV
K. V. OSSIAN DAHLGREN.
Redan under antiken hade man iakttagit, att det i mitten av mo rotens vita storflockar kunde sitta en mörkröd blomma (tig. 1). Färgen har sedan gammalt betecknats som purpureus, en poetisk beskrivning, som går igen hos kanske flertalet flor aförfattare. (Purpurröd och smaragdgrön äro som bekant tvenne ofta miss brukade prydnadsepitet.) Den i regeln svartröda, ja, för ögat ibland t. o. m. svarta blomfärgen har stundom beskrivits såsom »blodröd» ■— hos oss av Gertz (1906, s. 318) och Lindman (1905, s. 198) — men då måste man åtminstone tänka på levrat blod. I Gmelins flora (1805, s. 627) anges färgen vara »carmineus». Rafn (1800, s. 293) påstår, ehuru föga träffande, att ifrågavarande blommor skulle vara »kjodfarvede». I Neumans flora (1901) uppges, att mittflocken kan ha svart- eller rödbruna blommor. Buchenau (1894, s. 378) karakteriserar färgen som brunröd, och Schube(1904, s. 297) potentierar detta genom att tala om den såsom »dunkelrotbraun». 1 Karschs (1911, s. 132) llora över Westfalen anges mittblomman vara brun. Själv har jag verkligen också två eller tre gånger sett en blomflock med en tydligt brunfärgad mittblomma, men det var rena undantagsfall. Denna lilla exkursion i floralitteraturen må räcka; den visar ju hur skiftande omdömena ha varit. Ifrågavarande blommor äro normalt mörkröda, närmast svartröda (violettröda skriver v. Schwerin 1922, s. 144), men sällsynta gånger har också en ljusare röd eller rosafärgad blomma noterats.
I vår svenska floralitteratur är den underliga morotsblomman endast sparsamt omnämnd. Utom hos Neuman och Lindman (även 1918, s. 439) är den uppmärksammad av Areschoug(1881, s. 159), och den skymtar också svagt på en färgfotografi i Lagerbergs»Vilda växter i Norden», uppl. 2 (s. 1218, pl. 642). I den populära »Floran
MM J
: H '••f
w
Fig. 1. Daucus carola. Inflorescens med en ensam mörkröd mittblomma. Kantblom mornas yttre kronblad äro som vanligt större än de andra. — Något förminskad.
i färg» (Bolin & v. Post 1950, s. 179) omtalas att det »ofta sitter
en röd blomma i flockens mitt». I Kullenbergs (1957, pl. 4) ny
utkomna översikt av blombiologi i samband med insektsbesök lin nes en bild av en dylik Daucus-Üock, som visar dess svaga reflexion av ultraviolett ljus, och även samma blommor fotograferade i UV och synligt ljus (utan rött, som många insekter ej kunna uppfatta).
Den kalkhaltiga jordmånen på Öland tycks i hög grad gynna växten. (Där går den ofta under namnet »Drottningens spetsar», enligt vad laborator B. Kullenberg omtalat för mig.) Under ett par
somrar har jag vid mina besök på ön roat mig med att göra en del observationer över morotens egendomliga mittblommor (fig. 2). Ty värr hade det dock totalt fallit mig ur minnet, att redan Gertzför snart
tre decennier sedan hade skrivit en rätt utförlig rapport i Botaniska Notiser om sina iakttagelser över samma växt (Nyhamns läge, Skåne). Min första impuls var därför att låta mina egna anteckningar ligga oanvända, men jag har ångrat mig. De rätt instruktiva fotografier, som togos 1951 av till Uppsala hemfört levande material, förtjäna nog att återgivas; och Gertz’ framställning (1928) kan också kom pletteras med nya fakta, litteraturhänvisningar och synpunkter. För här ej citerade uppsatser hänvisas i huvudsak till sammanställ ningar hos honom, Penzig (1921, s. 429) och Hegi (1926, s. 1523).
583
»* *
Hg. 2. Daucus carota. Delar av tre inflorescenser, vardera med en ensam mörk mitt blomma.
I sin gradualavhandling meddelade Gertz (1906, s. 318) helt
kortfattat, att »den blodröda centralblomman i totalinflorescensen för anthocyan i perigonbladens epidermis». I sitt senare arbete hat- han dock bekräftat Möbius’ (1913, s. 324) uppgift om att även dessas inre celler skulle vara anthocyanförande, något som i hög grad skulle bidraga till att framkalla den mörka färgtonen. Senare har dock Küster (1942, s. 377) särskilt framhållit, att det just är
epidermiscellerna som »vorzugsweise oder ausschliesslich» äro färgade, medan själva grundvävnaden är färglös. (Hos rosenröda blommor skulle epidermis sakna eller endast ha obetydligt med anthocyan, medan däremot mesofyllet skulle vara relativt välförsett med färgämne, även om det kunde saknas i vissa skikt.) Några snitt, som i somras gjordes genom levande petaler, visade, att de något toppiga och svagt strierade epidermiscellerna voro fyllda av en stark färglösning som saknades hos mesofyllcellerna. Vid
Fig. 3. Daucus carota. Den ensamma mörka mitt blomman är mycket större än de andra blommorna och skjuter ofta upp över dem. — Först. 3:1.
tryckning av några andra blomblad befanns dock att anthocyan även kunde finnas i några inre mer eller mindre greniga celler; men framför allt är det epidermiscellerna, som äro färgade.
Som ett anmärkningsvärt förhållande måste omtalas, att de svart röda blommorna förblevo mörka, sedan de fixerats i en lösning med krom- och ättiksyra samt formol; och de ljusnade icke heller vid senare urtvättning i vatten, härdning genom spritskalan och över föring i butylalkohol för paraffininbäddning. Vid fixeringen måste väl någon substans ha uppkommit, som är olöslig i de nämnda vätskorna. Efter snittning med mikrotom visade sig epidermis cellerna innehålla en stor mörkfärgad kropp eller mera sällan två eller flera mindre kulor. Dessa torde väl motsvara de anthocyan- strukturer, som enligt Gertz utskiljas om cellerna bli utsatta för
tryck.
Den ofta påfallande stora centralblomman (fig. 3) måste be traktas som en reducerad småflock. Även en liten solitär ljusröd eller helt vit centralblomma går att leta upp. Någon enstaka gång kan reduktionen gå så långt, att endast en steril vävnadstapp är utbildad. Centralblommans ofta förhållandevis långa »skaft» bär i regeln två små motsatta svepeblad (ibland tre, någon gång flera eller bara ett enda). Att den röda centralblomman representerar en mycket redu cerad terminal småflock framgår särskilt tydligt av det mycket van liga förhållandet, att det i stället för den solitära blomman utvecklas en större eller mindre central partialinflorescens (fig. 4). Just som denna uppsats skulle sändas till tryckning, har Trolls (1957, fig.
307) nya Pflanzenmorphologie kommit hit, där han med en liten bildserie illustrerar hur den centrala röda småflocken kan reduceras till en enda blomma, som t. o. m. kan vara alldeles utan något svepeblad.
Sv. Bot. Tidskr., 51 (1957): i
Fig. 4. Daucus carota. Delar av tre inflorescenser som vardera ha en central småflock med mörkröda blommor.
Redan Warming har framhållit, att vi hos det Daucus tämligen
närstående släktet Artedia skulle ha ett slags motsvarighet till mo rotens röda centralblommor. Det utvecklas nämligen en starkt fär gad och förhållandevis tjock mittpelare, som avslutas med en massa mörkröda utskott, tätt besatta med korta hårbildningar. Det ligger nära till hands att tolka denna underlighet (fig. 5) som en ombildad terminalinflorescens med en mängd skaft utan blommor.
I en central småflock hos Daucus kunna ibland alla blommorna vara mörkröda. Två gånger räknade jag icke mindre än 10 sådana, och även talen 8-2 har jag noterat. Russell (1926, s. 611) har hos två centrala småflockar funnit 14 röda blommor i den ena och 17
*
%
W
.W/i
Fig. 5. Artedia squamata. Till vänster en hel inflorescens visande kantblommornas enormt stora yttre kronblad. I centrum sitter den egendomliga »mittpelaren*. — Till höger det förstorade centrala partiet av samma blomställning, där man tydligare ser »mittpelaren» med dess buske av röda, korthåriga utskott. (Herb. Upsal. Jericho 1912.
S. Linder leg.)
i den andra. I en stor central partialinflorescens är det dock mycket vanligt, att en del av blommorna äro vita. I ett extremt fall sågos 9 röda och en ensam vit blomma; i ett annat två likstora blommor, den ena röd och den andra vit. Ofta finnas en, ett par eller flera större mörkröda blommor, som till grannar ha mindre sådana av samma slag men därjämte också helt vita blommor. Det är också synnerligen vanligt, att utom de rent vita blommorna även finnas sådana som äro sektorialt uppdelade eller fläckade i rött och vitt. (Även sådana fall ha t. ex. antecknats som 7 helröda blommor jämte en blomma med tre vita kronblad och en annan gång 3 helröda och en med två vita blad.) Någon gång kan också en ensam stor central blomma vara både röd och vit. Jag har sålunda observerat vita marginalpartier på något eller några av kronbladen. En gång note rades att två av petalerna voro vita, och en annan gång att precis hälften av blomman var vit, dvs. att ett av de fem bladen hade en vit och en röd halva. Gränsen mellan de båda färgerna är skarp, och man letar förgäves efter övergångszoner med så småningom uttunnande färgnyanser. Man kan dock någon gång få se ljusröda fläckar. I en rikblommig centralflock befunnos 2 blommor vara mörkröda, 5 både mörkröda och vita, 2 ljusare röda, 3 rosafärgade och vita samt 1 både mörkröd, rosafärgad och rent vit. Kanske det var fråga om olika stadier av färgutvecklingen. Som nämnt har
Fig. 6. Daucus carota. Utom den centrala småflocken resp. mittblomman ha även några av blommorna i en grannflock blivit mörkröda. •— Först. 2:1.
jag också några gånger sett en liten ljusröd centralblomma och även en rätt stor sådan. Möbius (1. c., Tal. I, lig. 10) har också
avbildat en dylik.
Icke sällan har utom den solitära centralblomman eller den cen trala delinflorescensen också en liten grannflock — även tre dylika har jag sett — röda blommor eller både röda, partiellt röda och helt vita sådana (fig. 6). Ibland kunna dessa småflockar vara förenade, särskilt nedtill. Även Russell (1926, s. 612) har flera gånger iakt tagit delta, och en gång har han sett tre hopvuxna partialinflore- scenser. Stundom kunna som nämnt två småflockar i mitten av blomställningen ha idel röda blommor; men i allmänhet finner man åtminstone i den ena av dem vita eller spräckliga blommor. Det bör framhållas att de röda blommorna i den ena flocken då all tid vetta mot de röda (eller den ensamma röda) i den andra (fig. 6).
Man har beskrivit en var. atropurpurea (»floribus omnibus atro- purpureis»), där samtliga blommor skulle vara mörkröda. Huru vida detta genomgående är fallet är väl en smula osäkert.* 1 Den bekante Genéve-botanisten Briquet (1922) har sålunda studerat en
intressant population på ett ställe nära St Paul i Savojen. Bland
tal-1 En f. atrombens Lge har hos oss observerats i Landskrona-trakten av Nilsson
(1952, s. 87), som skriver: »Denna mycket mörkt brunröda form av vildmorot har under senare år iakttagits i talrika ex. i strandplanteringarna vid Erikstorp. I Landskrona- trakten äro färgade former förut anmärkta från Örja (Sylvén 1931 b).» Gotländska exemplar med måttligt starkt rödfärgade blommor finnas i Uppsala-herbariet. På Öland har jag ofta sett individer med rosafärgade (eller åtminstone i början ljusröda) i stället för vita blommor, men dessa exemplar tyckas dock icke ha större benägenhet än andra för att utveckla de mörkröda centralblommorna.
* * t * ,* .**% * wfL ?V-*
* * v #
t
Fig. 7. Daucus carota. En hel sektor av inflorescensen har blommor av samma mörk röda färg, som ofta utmärker den centrala småflocken. — Något förminskad bild
efter Patch.
rika normala Z)aucizs-plantor uppträdde där åtskilliga, som visade en stark »mélanérythrisme». Han kunde dock icke upptäcka något exemplar, vars alla flockar voro mörkröda. I själva verket kunde alla övergångar påträffas mellan flockar med en ensam svartröd mittblomma och sådana, som åtminstone praktiskt taget hade enbart mörka blommor; och förhållandena kunde vara högst växlande hos ett och samma individ. Vi kunna här också anföra ett par för Briquet okända amerikanska meddelanden om en dylik utbredd melan- erythrism. År 1919 rapporterade Robinson (s. 70) om fyndet av ett exemplar, där nära hälften av en storflock från centrum till peri ferien hade fått samma mörka färg som mittblomman, även om en del av blommorna delvis hade vita petaler eller röda sådana »striped with white». Samma år omtalade också Eames (1919, s. 147), att han sett »a completely colored specimen». Även rosafärgade flockar har Eames iakttagit liksom intermediära fall, där alla eller en del av petalerna i somliga marginalblommor kunde vara mörkröda, en färg som mot centrum vanligen gradvis övergick till rosa eller rent vitt. En intressant Daucus-flock har senare fotograferats av miss
Edith Patch i U.S.A. och publicerats i Journal of Heredity (1930, s. 386). Då en sektor av storflocken här hade idel svartröda blommor (fig. 7), kommer man givetvis först att tänka på en vegetativ mutation; men särskilt med kännedom om Briquets iakttagelser är väl en sådan i alla fall ganska dubiös. Det är ju fråga om en långt i sidled utstrålande färgning, som normalt endast brukar uppträda i blom ställningens centrum.
De stora petalerna hos den mörkröda centralblomman, som ofta inte öppnar sig, äro förhållandevis styva, förbli gärna hopvikta och inåtböjda samt persistera länge. Blomman visar ofta oregelbunden heter i sina talförhållanden, och särskilt gäller detta, såsom man ofta har iakttagit, gyneceet. Detta är icke sällan tretaligt. Tydligen är den stora blomman ofta uppkommen av två eller flera samman smälta blomanlag, ett synanthium alltså. Gyneceet har jag också sett vara sammansatt av 4, 6, 7 och 8 karpeller. I de båda sistnämnda fallen voro blommorna tydligen uppbyggda av två komponenter, vardera med resp. 3+4 och 4 + 4 fruktblad. — En uppdelning i röda och icke röda partier kan vara mycket påfallande i central blommans gyneceum. Här kan ofta både disken (stylopodium) och stiften vara röda; men det finnes också vit disk och röda stift eller, fast sällan, omvänt. Både disken och stiften kunna även vara vita. Ståndarna äro antingen röda eller vita; och samma blomma kan någon gång härbärgera båda slagen.
Särskilt i flororna uppges ibland att de mörka centralblommorna skulle vara sterila. »Golde» står det sålunda hos Blytt(1906, s. 541),
Nordhagen (1940, s. 467) och Lange (1851, s. 183 och i följande upplagor) för att nu till att börja med endast citera några nordiska författare. »Flocculus centralis sterilis» uppger Gmelin(1805, s. 627), och »Flore umbellae centrali typice sterili, atropurpureo» läsa vi i en flora över Spanien (Willkomm & Lange 1880, s. 21); steril skriva
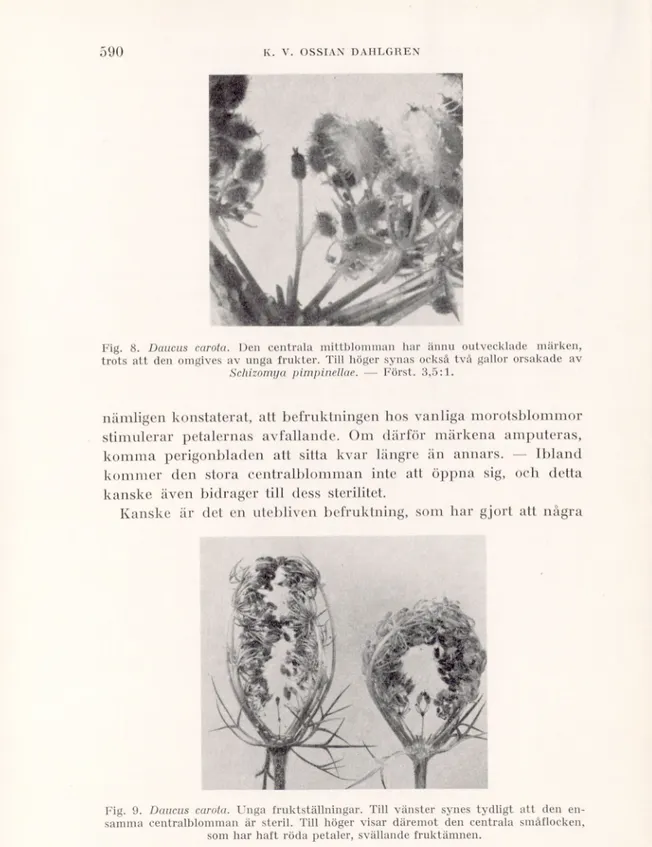
Coste (1903, s. 157), Musciiler (1912, s. 713) och v. Schwerin (1922, s. 144); »neutral» står det i Smiths gamla flora (1824, s. 39); »unfruchtbar» uppgiva Gradmann (1900, s. 255), Schinz & Keller (1923, s. 504) samt flera andra. Det anförda får räcka. Blommorna i Iråga bli otvivelaktigt ofta ofruktsamma (lig. 8 och lig. 9, t. v.). Deras stora styva kronblad kunna ibland sitta kvar, sedan de unga fruktsamlingarna börjat sluta sig, och kanske detta åtminstone till en del kan sammanhänga med att befruktning har uteblivit. Tre amerikanska forskare (Borthwick, Phillips & Robbins 1931) ha
Klr
t t
Fig. 8. Daucus carota. Den centrala mittblomman har ännu outvecklade märken, trots att den omgives av unga frukter. Till höger synas också två gallor orsakade av
Schizomya pimpinellae. — Först. 3,5:1.
nämligen konstaterat, att befruktningen hos vanliga morotsblommor stimulerar petalernas avfallande. Om därför märkena amputeras, komma perigonbladen att sitta kvar längre än annars. — Ibland kommer den stora centralblomman inte att öppna sig, och detta kanske även bidrager till dess sterilitet.
Kanske är det en utebliven befruktning, som har gjort att några
Fig. 9. Daucus carota. Unga fruktställningar. Till vänster synes tydligt att den en samma centralblomman är steril. Till höger visar däremot den centrala småflocken,
som har haft röda petaler, svällande fruktämnen.
Fig. 10. Daucus carota. Den ensamma centralblomman har av gallmyggan Schizomya
pimpinellae förvandlats till en egenartad fruktliknande bildning. (Jfr fig. 8 t. h.!) —
Först. 3,5:1. ■É jj
t* m J*fiT
S + \/ > i
5» ■? _ g t * if %författare ha betecknat den mörka centralblomman som »verküm mert» (Gilg & Schürhoff 1931, s. 301) eller uppgivit att den åt
minstone i allmänhet är förkrympt (Buchenau 1894, s. 378; Karsch
1911, s. 132). »Outvecklad» skriver också Aresciioug(1881, s. 159).
1 Bonniersstora och praktfulla floraverk (1921, s. 80) uppges egen
domligt nog, att den purpurfärgade centralblomman skulle sakna såväl pistill som ståndare; men det är då en smula motsägande att längre ned, i redogörelsen för olika slags anomalier, läsa, att denna blomma kan ha 3-8 karpeller. Mera roande än riktigt är det be fängda påståendet i Karstens flora (1895, s. 419): »... mittleres
Döldchen oft in eine purpurne Drüse verändert». Att fruktsättning verkligen kan inträffa är dock fastställt. Vollman (1914, s. 574)
skriver visserligen endast, att blomman är »meist unfruchtbar», och
Garcke (1922, s. 570) t. ex. säger »oft unfruchtbar»; men »meist
fruchtbar» står det helt positivt hos Nicolaisen (1950, s. 221), och i
sitt korta föredragsreferat betecknade Kronfeld (1892, s. 12) blom
man i fråga kort och gott som »fruchtbar». Russell (1926, s. 614)
avslutar sin rapport från Frankrike med att framhålla, att sterilitet visserligen är vanlig, men att han i slutet av säsongen dock har stött på några frukter, som sutto isolerade i centrum och hade »la colo ration rouge caractéristique des fleurs anormales du D. Carota».
Briquet och sedan Gertz ha påpekat, att redan Darwin obser
verade fruktsättning. I fig. 9 till höger synes en central småflock med svällande fruktämnen. Hos de röda centralblommorna konstaterade
Håkansson (1923, s. 84) alldeles normala embryosäckar; och själv
har jag iakttagit detsamma. I sin utförliga beskrivning av de embryo logiska förhållandena nämner Bortiiwick (1931) ingenting om dessa
blommor, vilket väl beror på att han endast har studerat kultur former av morotsväxten.
I förbigående kan här också omnämnas att tydligen även själva centralblomman kan omvandlas lill det säregna cecidium (Boss 1911, Taf. III, fig. 47), som orsakas av gallmyggan Schizomya pim- pinellae. Jag har ett par gånger konstaterat detta (fig. 10). Ur de förhållandevis stora violett anlupna fruktgallorna av denna typ kan man långt före Ilockens fruktsättning peta fram de vackert orange- färgade larverna. (Inom parentes må här också noteras att Florence
Flemion och medarbetare 1949, s. 299, ha meddelat, att en annan
insekt, Lygus oblineatus, kan hos dill och Daucus förstöra embryo- nerna under så gott som alla utvecklingsstadier men likväl lämna endospermet lill synes alldeles oskadat, varför till det yttre normala men icke grobara frön kunna bildas.)
I ett par arbeten har jag sett bestämda uppgifter om att röda central blommor alltid skulle utvecklas. De finnas »regelmässig», skriva sålunda Gilg & Schürhoff (1931, s. 301) i sin lärobok, och »stets an allen von uns untersuchten Exemplaren» står det i Röh- lingsgamla flora (1826, s. 359). Många floror uppgiva endast, att i flockens mitt finnes en röd blomma, men gärna är ordet »ofta» eller »i allmänhet» inskjutet. I själva verket är det nämligen visst icke alla Daucus-inflorescenser, som utveckla röda mittblommor. »Gjen- nemsnitlig hver tiende» säger Warming (1876, s. 97). Andra tal citeras av Gertz (1928, s. 311, not) och Küster (1. c., s. 373, not). Eiiinger (1923, s. 180) fann i Schweiz, att 22-45% av plantorna kunde i somliga Ilockar ha och i andra samtidigt sakna de röda mittblommorna. Dessa uppgifter stå dock i rätt stark kontrast till vad jag själv har observerat på Öland. Enligt min erfarenhet är det allra vanligast, att de mörkröda blommorna antingen finnas eller också saknas hos ett individs alla blomställningar. Mången gång kan det naturligtvis vara svårt att vid ett visst tillfälle göra ett be stämt uttalande, då en del flockar kunna befinna sig på ett så tidigt utvecklingsstadium, att man icke vågar säga hur de senare komma
593 att gestalta sig. På en torräng och längs landsvägskanter undersökte jag sammanlagt 1183 exemplar. Av dessa hade 471 röda blommor i alla sina flockar, 597 saknade lika genomgående dylika, och hos 115, dvs. 9,72%, kunde man finna blomställningar av båda typerna; oftast var det då blott en eller annan flock som avvek ifrån de andra. (I materialet ingingo även en del små exemplar, som blott hade en utvecklad flock.)
Hos odlade morötter har jag aldrig sett de egendomliga central blommorna; och enligt benäget meddelande från A.B. Ölandsfrö, Mörbylånga, har man där i sina stora fröodlingar av sorterna Nantes, London torg och Flakkeer icke heller observerat de röda blommorna.
Nicolaisen (1950, s. 231), som behandlat moroten i Roemers och
Rudolfs »Handbuch der Pflanzenzüchtung», har ej heller hos kultur
former sett någon »Mohrenblüte», såsom tyskarna med en ordlek gärna kalla den mörka mittblomman; men han citerar Becker, som har uppgivit att förekomsten av en sådan blomma skulle vara myc ket växlande hos kultursorter. I Gmelins förut omnämnda flora
uppges, att de röda centralblommorna ofta uppträda »in individuis spontaneis et cultis». Kanske detta kan sammanhänga med inkors- ningar av vild morot? Deichmann (1890, s. 77) har fäst uppmärk
samheten på att hybridisering med vild morot är ganska vanlig och att detta kan inverka ofördelaktigt på kultursorternas avkomma. I brev har fil. dr H. Lamprecht, Weibullsholm, nu också meddelat, att under vissa år och i vissa sorter kan i fröodlingar en och annan planta utveckla röda centralblommor; men en undersökning har då alltid visat, att detta berodde på korsning med vild morot. Baur
(1924, s. 3, not) har även omnämnt, att den vilda typen kan bli befruktad av den odlade.
Somliga Daucus-arter kunna utveckla och andra sakna de röda centralblommorna. I Costes flora (1903) över Frankrike och när
gränsande områden upptagas åtta arter, och av dessa uppges fyra ha och fyra sakna »une fleur purpurine». Det ligger nära till hands att förmoda, att även hos vår vilda morot genetiska olikheter göra vissa individer mera benägna än andra att utveckla röda central- blommor. På åtskilliga tyska ståndorter äro de »mohrblütige» säll synta eller saknas alldeles (Küster 1942, s. 373). Det uppges så
lunda, att man på de Ostfriesiska öarna på flera ställen förgäves får leta efter exemplar med dylika blommor. Goebel (1928, s. 552)
tycks anse, att uppkomsten av dessa på något sätt skulle samman hänga med en plantas allmänna utveckling, ty »wenigstens bei
kräftigen Exemplaren» säger han sig ha sett blomtypen i fråga. Ut med vägkanter på Öland har förf. flera gånger sett luxurierande plantor — med ända till ett tjog och ännu flera blomflockar utveck- lade —, som växte bredvid varandra, men där ändock det ena exemplaret genomgående saknade, medan det andra lika konse kvent hade utvecklat de röda blommorna. I populationer av Dauern cavota såg jag ibland individer, där särskilt de perifera blommorna voro svagt rosafärgade, åtminstone i början. Redan på ett mycket tidigt stadium kunde då flockens knoppar vara rödaktigt anlupna, fast de utslagna blommorna sedan i många fall tycktes bli lika vita som hos andra exemplar; men fruktämnena och småflockarnas skaft brukade då icke vara rent gröna utan stöta i en mörkare färg ton. Dylika plantor — liksom sådana med permanent rosafärgade blommor — hade icke större benägenhet än andra för att utveckla röda centralblommor, såsom det förut har påpekats i en not.
En noggrann beskrivning av blomställningens utveckling hos od lad morot har lämnats av tre amerikaner (Borthwick, Phillips & Robbins 1931). Umbellaternas inflorescenser höra till de »obegrän
sade» blomställningarna. Hos sådana kan understundom det ter minala vävnadspartiet förbrukas för att alstra en toppblomma, som sålunda begränsar den till sin typ racemösa blomsamlingen. En dylik blomma får ibland en från de andra blommorna avvikande byggnad. Vi'behöva endast erinra om den stora aktinomorfa topp blomman hos vissa Digit alis-stsunmar. Hos blåklockor slår ofta en terminal blomma ut tidigare, trots sin senare anläggning, än de knoppar, som sitta närmast i klasen (Goebel 1931, s. 60).1 Såsom
man har framhållit är också den stora, luxurierande mittblomman resp. partialinflorescensen hos Daucus att anse såsom gynnad, »ge fördert». Särskilt hos vissa individer tycks dock det meristematiska cellmaterialet i blomställningsanlagen så att säga icke räcka till för att frambringa den stora mittblomman eller en terminal, gärna mer eller mindre röd småflock. Detta liksom de teratologiska avvikelserna
1 I anslutning till en tidigare uppsats (Dahlgren 1943, s. 403) kan förf. i förbi gående också omnämna, att han nere vid Kalmarsund, Vickleby socken, Öland, år 1951 såg ett exemplar av Campanula rotundifolia, som hade ett skott med åtta nor mala blomknoppar men vilket avslutades med ett stort synanthium, sammansatt av fem stycken blommor. Den vida, grunda och något veckade blomskålen visade 20 likformiga kronbladsflikar och omgavs av 20 stycken foderblad. Även ståndarna voro 20 (två med avfallna knappar), och 15 märkesflikar kunde räknas.
hos de stora mörka blommorna talar ju också för att deras yppighet närmast vore en extravagans, som framkallats av speciellt »gynn samma» förhållanden under den tidiga utvecklingen.
Den som bekant tvååriga morotsplantan fordrar en tids kyla för att kunna lämna sitt rosettstadium, skjuta upp och övergå till blombild ning. Genom behandling med det under de senaste åren så omtalade gibberellinet kunde dock Lang (1957, s. 711) framtvinga blomning
utan någon föregående köldbehandling hos morot och några andra långdagsväxter. Rubbningar av hormonal karaktär kanske också betinga centralblommornas avvikande morfologi hos den vilda mo roten. Hos Saponaria officinalis kunde Monique Astré (1957, s. 933)
framkalla allehanda anomalier, bl. a. övertaliga karpeller, genom inverkan av 2.4-diklorfenoxiättiksyran. Att iögonfallande modifika tioner kunna uppkomma i samband med ändrade närings- och hormoninflytanden, framgår också av vissa ympningsförsök för att nu inte tala om cecidiologiens vittnesbörd. Nyligen har Mathilde
Pichenor (1957, s. 1669) under sina solanacéstudier t. ex. visat,
att de skott av Petunia violacea, som ympades på Datura stramonium, växte yppigare och buro större blad än normalt. De ovanligt stora blommorna (6,3 cm mot annars 4,2) slogo ut senare, fmgo en annan färg och utbildade större frukter med c:a 100 fler frön (därav dock 27% missbildade) än kontrollplantorna.
Vad nu den röda färgen beträffar lämnade Nelson (1954, s. 143)
för ett par år sedan ett intressant meddelande. I Positano vid Salerno- viken upptäckte han en Daucus-population, där samma slags taggar som på fruktämnet också utvecklades på de röda blommornas petaler men däremot aldrig på de vitas. Om flera röda blommor framkommo, sutto dessa taggar tätast på blommorna närmast själva centrum. Mera vid sidan därav kunde taggar t. o. m. alldeles sak nas, trots att kronbladen voro röda. Han ser i den större eller mindre benägenheten att utveckla taggar en direkt parallellföreteelse till anthocyankoncentrationen, som i sin tur skulle vara avhängig av blommornas placering. Även om vårt öga icke skulle kunna märka någon skillnad i färgintensitet mellan olika blommor, menar han dock, att en sådan skulle finnas och att taggarnas mängd skulle vara ett slags indikator på kronbladens anthocyanhalt. Relata refero utan kommentarer.
Nyligen ha två forskare, Bayer & Wegmann (1957, s. 37), påvisat
ett enzym, cyaninoxidas, vilket med o-fenoler som aktivatorer över för syre till anthocyanfärgämnen och därigenom kommer den röda
färgen att försvinna. Många unga växtdelar äro som bekant först röda, fast denna färg så småningom plånas ut. De förut omtalade rosafärgade blommorna hos Daucus t. ex. vitnade i tlera fall senare. Man kan föreställa sig, att centralblommans anthocyanrikedom skulle kunna bero antingen på att enzymet i fråga upphörde att produceras eller också på att de aktiverande orthofenolerna icke längre bildades. (På samma sätt skulle, såsom de båda författarna framhålla, vid mutationer s. k. blodbokar, blodlönnar etc. kunna komma till stånd och likaså den höstfärgning av bladen, vilken betingas av anthocyan.)
Som förut har framhållits synas under vissa betingelser en del men icke alla individer och raser ha en genetiskt grundad förmåga att i blomställningens mitt utveckla de mörkröda blommorna. För anthocyanbildningen är socker nödvändigt, och man kunde därför vara benägen att också sätta blommornas starka färg i samband med en särskilt riklig tillströmning av socker. [Schröder (1934,
s. 717) har i sina försök med Petunia — till vilka vi skola återkomma — visat, att en extra sockertillförsel kunde öka anthocyanmängden i cellerna.] Goebel (1928, s. 552) framhåller särskilt detta. Om
flera blommor utvecklas i den centrala flocken, uppger han, att de skulle bli mindre och dessutom svagare färgade, men detta stämmer dock icke alltid. På skott, som utvecklades från avbetade Daucus- exemplar, har Molliard (1921) — utom ett flertal missbildningar —
iakttagit, att karpellerna ibland blevo röda och »communiquent å Pinflorescence une teinte tres anormale» (s. 474), vilket allt han förmodar orsakas av förändringar i näringsmiljön. Gertz (1928,
s. 312) menar, att färgämnets fördelning i blommorna »får till skrivas komplikationer i byggnaden och förloppet av de till dem ledande kärlknippena». Genom ledningsbanornas olika härkomst och deras anastomoser förmodar han, att man får en förklaring till att vita blommor kunna sitta invid röda och till att blommorna ibland endast delvis äro färgade. Det ligger i viss mån något bestickande i
Goebels och Gertz’ så att säga rent anatomiskt-mekaniska syn punkt på fenomenet: en lokalt riklig tillförsel av socker skulle kunna föranleda en extra stark anthocyanfärgning; men vi få kanske nu också tillägga: i samband med rubbningar av den normala balansen mellan enzym och coenzym. Uppkomsten av en storflock med en mörkröd sektor (fig. 7), liksom över huvud taget en allmännare röd- färgning av inflorescensen, skulle på så sätt också kunna få en någorlunda nöjaktig förklaring.
De skarpa gränserna mellan vitt och rött liksom kronbladens starkt varierande färgmosaiker i de delvis röda partialinflorescen- serna synas mig dock — liksom även den växlande färgfördelningen i den mörka centralblommans gyneceum — vara mycket svåra att förklara såsom endast beroende på förändringar av den normala ämnestillförseln. Förutom det genetiska underlaget måste man nog tänka sig, att också någon annan faktor eller andra faktorer på ett tidigt stadium äro av betydelse för den kommande utvecklingen. Härför tala de betydande förändringar av färgning och färgmönster, som man experimentellt har kunnat framkalla hos flera växter och särskilt hos en ingående studerad form av en Petunia hybrida. Genom undersökningar av Schröder (1934) och andra har det visats, att
om skott av denna utsättas för hög temperatur eller om de endast få en svag eller kortvarig belysning, påverkas de unga knopparna så, att blommorna sedan bli helt violetta. Om man däremot experi menterar vid en förhållandevis låg temperatur och i starkt ljus, kunna blommorna bli helvita. Genom olika gradationer av temperatur- och belysningsförhållandena kan ett exemplar tvingas att utveckla blom mor, som framvisa alla möjliga mellanstadier mellan den helt vio letta och den helt vita typen. Det brokiga utslaget av de växlande väderleksförhållandena torde nog också många ha observerat på de Pefimia-plantor, som bruka pryda våra balkongträdgårdar. Det är, väl att märka, själva färgmönstret, som varierar; och det förekom mer icke någon kontinuerlig övergång mellan vitt eller violett genom att cellernas anthocyanhalt gradvis minskar eller ökar. Gränserna mellan färgade och ofärgade områden är alltid skarp. En svagare belysning kan visserligen resultera i en svagare anthocyanfärgning, men detta inverkar icke på själva färgmönstrets beskaffenhet. Detta och färgintensiteten (anthocyanhalten) äro icke beroende av var andra. Den färgfördelning, som en gång har inducerats då de unga knopparna blott voro 1—2,5 mm långa, blir bestående hur än sedan de yttre retningsfaktorerna komma att växla. Det är sålunda endast under en helt kort period, en period som knappast torde överskrida 12 timmar, som knopparna kunna påverkas. — Då det som nämnt inte utbildas någon övergångszon mellan röda och vita områden hos de centrala morotsblommorna, är det frestande att förmoda, att även här växlande väderleksförhållanden under tidiga utvecklingsstadier kunna inverka på blommornas blivande färgmönster. En prövning av denna arbetshypotes är av flera skäl svår för mig att genomföra; men jag tror dock att detta förslag till en förklaring förtjänar att bli
framlagt, då det kanske kan stimulera någon ung forskare till att experimentellt pröva uppslaget.
Vi avstå från att här redogöra för några delvis rätt naiva teleolo- giska utläggningar (färgen skulle vara ett lockmedel för somliga djur och verka avskräckande på andra, däribland getter) samt ett par andra funderingar (nedärvd gallbildning och atavism), som de mörka mittblommorna ha givit anledning till. Dessa spekulationer ha i huvudsak numera endast kuriositetsintresse. På Öland vältra sig gärna ett slags vackra gula skalbaggar, Cteniopus flavus, i de stora blomflockarna; och de, liksom det fåtal andra insekter, vilkas blom besök jag mera flyktigt har iakttagit, lockas väl varken direkt dit eller repelleras därifrån genom närvaron eller frånvaron av de mörkröda blommorna i mitten. Naturligtvis böra dock insekter kunna uppfatta kontrasten mellan dessa och allt det vita. Vännen
Kullenberg har också meddelat mig, att han en gång råkade att
filma den vanliga guldbaggen (Cetonia aurata) just som den kröp fram och bet i den mörka mittblomman. Denna har också upp- givits (Hansgirg 1893, s. 262) verka tilldragande på vissa insekter
genom sin för oss obehagliga lukt. Tio personer, som jag lät »nosa» på en liten hög med 150 mörkröda blommor och en annan med vanliga vita, kunde icke märka någon särskild odör (ett par för- nummo över huvud taget endast en mycket svag doft) från de röda blommorna, medan de vita däremot föreföllo några av försöks personerna att ha en starkare doft. Inte heller Detto (1905, s. 327)
har kunnat känna någon speciellt motbjudande lukt från tie s. k. Mohrenblüten.
Summary.
The dark red central flowers of the wild Daucus carota.
Ever since ancient times it has been known that a dark red flower may appear in the centre of the inflorescence of Daucus carota (Figs. 1 and 2). This has been described or at least mentioned in quite a number of works. In the flora manuals that actually mention the divergent flower its colour is generally described as purpureus or atropurpureus, but terms such as sanguine, carmine, magenta, violet- red and flesh-coloured have also been used. Normally the colour is dark red, at times it even gives the impression of being black. Occasionally the flower in question may be a lighter red, distinctly brown or altogether white.
At first Gertz maintained that the anthocyan of the flower was
situated in the epidermis of the perigonal leaves, but in a later work he supported Möbius, who had stated that the cells of the paren chymatous tissue were also anthocyan carriers. Later, however,
Küsterreported that it was preferably or exclusively the cells of the
epidermis that were coloured, whereas those in the mesophyll were colourless on the whole. The present author has found that the cells of the epidermis are sometimes the only anthocyan carriers, but has also observed that some faintly branched mesophyll cells may be red.
After the dark red flowers had been killed in a mixture of formal dehyde, chromic and acetic acid, and had subsequently been treated with alcohol, the petals were still dark coloured, indicating that a sub stance nonsoluble in alcohol had been formed.
1 he solitary central flower (Fig. 3), which is often unusually large, must be regarded as a reduced terminal umbellet. This may be concluded not only from the involucral bracts but also from the fact that it is frequently replaced by a larger or smaller umbellet, whose flowers are at least partly dark (Fig. 4). A sort of parallel to the red central flowers of the carrot is found in the relatively related genus Artedia, which develops a coloured central pillar bearing a mass of dark red excrescences thickly covered with short trichomas (Fig. 5). This easily suggests the interpretation that this organization is a transformed and sterile umbellet.
Apart from the red flowers white ones are also not uncommonly found in the central umbellet of the wild carrot. Moreover flowers also occur that are divided into sections or spotted red and white. Even a large, single central flower may sometimes have pure white parts. Occasionally a small, neighbouring umbellet may have dark red flowers alone or in combination with white flowers. The red flowers in the one umbellet always turn towards the red ones in the other (Fig. 6).
There is a description of a var. atropurpurea where all the flowers are said to have the dark red colour. It is a little doubtful whether this is always so, especially in the light of Briquet’sinvestigations.
In the population studied by him there were umbels with every transition from those with a single red central flower to those with practically only dark red flowers. The circumstances could differ greatly in one and the same individual. Fig. 7 shows an umbel with a sector of purely dark red flowers, but it is improbable that this is a case of vegetative mutation.
The petals of the dark red central flower are relatively stiff, long remain folded up and bent inwards, and persist for a long time. The flowers frequently show irregularities in their numerical arrange ments, this being especially true of the gynoeceum. The latter is often threefold, but the author has also seen 4, 6, 7 and 8 carpels. Synanthia are apparently common. Sometimes the stylopod and the styles are red or white, while a white stylopod with red styles and vice versa may occur.
Especially in floral literature it is often stated that the red central flowers are sterile. The embryo sac, however, is normally developed, and though sterility is no doubt very common (Figs. 8 and 9 left) fruit setting can take place (Fig. 9 right). As previously mentioned the red perigonal leaves often persist for a long time, possibly owing to the fact that fertilization has not taken place. The latter process is known to stimulate the shedding of the petals in the cultivated carrot flower (Borthwick et al.).
The solitary central flower, too, may sometimes be attacked by the gall insect, Schizomya pimpinellae, and thus be transformed into a curious cecidium (Fig. 10).
Red central flowers are not always found. Specimens with and without them may be found side by side. According to the author’s observations on the island of Öland in the Baltic all the inflores cences of one individual usually either develop or do not develop the red flowers, indicating genotypical differences. The author once counted 471 specimens with red flowers in all the umbels, 597 with only white ones and 115 (9.72%) with inflorescences of both types. Specimens with pink flowers are sometimes seen, at least in the beginning, but these plants show no greater tendency than others to develop dark red central flowers.
In the cultivated carrot the author has not observed the red cen tral flowers. A few reports of the appearance of such flowers are likely to be connected with bastardization with the wild carrot.
Bayer & Wegmann (1957) have recently demonstrated the occur
rence of an enzyme, cyanin-oxidase, which breaks down anthocyan when it is activated by orthophenols. It is therefore not impossible that the red central flowers as well as an extensive redness of the inflorescence may be due either to the cessation of enzyme produc tion or to the circumstance that coenzymes are no longer formed. Sugar is needed for the anthocyan synthesis, and one may therefore also find a link between the dark red colour of the central flowers
and an especially abundant sugar supply (Goebel), just as there is reason to suppose that their luxuriance is usually connected with a relatively generous food supply. Goebel and Gertz think that
the course of the vascular strands sometimes causes some of the flowers in the centre to be red and others to be white, and that some are sectionally coloured red and white. This simple mechanical- anatomical theory is undoubtedly attractive, but conditions are probably more complicated.
The significant changes in the colour patterns that can be pro duced in Petunia hybrida and other plants by exposing very young buds to varying conditions of temperature and light support the suggestion that changing weather conditions may also have an in fluence on the distribution of red colouring matter in the central flowers of Daucus. Just as in Petunia, where the early induced extension of violet and white sectors is sharply defined, so Daucus, too, lacks transitional zones between the red and white parts. The author hopes that the working hypothesis stated above may inspire someone to attempt an experimental verification.
Uppsala, Institutionen för systematisk botanik, hösten 1957.
CITERAD LITTERATUR.
Areschoug, F. W. C., Skånes flora innefattande de fanerogama och orm- bunksartade växterna. Uppl. 2. — Lund 1881.
Astré, Monique, Induction de pieces surnuméraires et de caractéres »d’inflorescences» chez Saponaria officinalis L. —• C. R. hebd. d. séances de l’Acad. d. Sei., 244. Paris 1957.
Baur, E., Die wissenschaftlichen Grundlagen der Pflanzenzüchtung, ein Lehrbuch für Landwirte, Gärtner und Forstleute. Aufl. 3-5. — Berlin 1924.
Bayer, E., & Wegmann, K., Enzymatischer Abbau von Anthocyanen. - Zeitschrift f. Naturforschung, 12 b. 1957.
Becker, J., Handbuch des Gemüsebaues. Aufl. 2. — Berlin 1929. (Cit. efter Nicolaisen.)
Blytt, A., Flaandbog i Norges Flora. — Kristiania 1906.
Bonnier, G., Flore complete illustrée en couleurs de France, Suisse et Belgique, 4. — Paris 1921? (utan tryckår).
Bolin, L., & v. Post, L. O. A., Floran i färg. Uppl. 2. — Stockholm 1950.
Borthwick, H. A., Development of the macrogametophyte and embryo of Daucus carota. — Bot. Gaz., 92. 1931.
Borthwick, H. A., Phillips, Mabel, & Robbins, W. W., Floral develop ment in Dauern carota. — American Journ. of Bot., 18. 1931.
Briquet, J., Le mélanérythrisme floral chez le Daucus Carola. — Annuaire du Conserv. et du Jard. bot. de Geneve, 21. 1922.
Buchenau, F., Flora der nordwestdeutschen Tieflandsebene. — Leipzig 1894.
Coste, H., Flore descriptive et illustrée de la France, de la Corse et des contrées limitrophes, 2. — Paris 1903.
Dahlgren, K. V. O., Om olika slags dubbla blommor hos blåklockor, särskilt hos Campanula rapunculoides. — Svensk Bot. Tidskrift, 37. 1943.
Deichmann, A. W., Krydsbefrugtning hos Gulerodder. — Om Landbrugets Kulturplanter. No 8. 1890. (Blott ref. sett i Bot. Centralblatt, 49, s. 271. 1892.)
Detto, C., Blütenbiologische Untersuchungen. I. — Flora, 94. 1905.
Eames, E. H., Another exceptional specimen of Daucus Carota. — Rhodora, 21. 1919.
Ehinger, M., Notes relatives au fleuron pourpre des ombelles du Daucus Carota L. — Bull, de la Soc. Vaudoise d. sei. nat., 55. 1923.
Flemion, Florence, Poole, Harriet, & Olson, June, Relation of Lygus bugs to embryoless seeds in Dill. — Contrib. fr. Boyce Thompson
Institute, 15. 1949.
Garcke, A., Illustrierte Flora von Deutschland. Aufl. 22. —- Berlin 1922.
Gertz, O., Studier öfver Anthocyan. — Diss. Lund 1906.
—»—, Om anthocyanblomman hos Daucus Carota L. — Bot. Notiser 1928.
Gilg, E., & Schürhoff, P. N., Grundzüge der Botanik für den Hoch schulunterricht. — Breslau 1931.
Gmelin, C. C., Flora Badensis Alsatica etc., I. — Carlsruhe 1805.
Goebel, K., Organographie der Pflanzen insbesondere der Archegoniaten und Samenpflanzen. Erster Teil. Aufl. 3. ■— Jena 1928.
—»—, Blütenbildung und Sprossgestaltung (Anthokladien und Inflores zenzen). Zweiter Ergänzungsbd. z. Organographie d. Pflanzen. - Jena 1931.
Gradmann, R., Das Pflanzenleben der Schwäbischen Alb mit Berück sichtigung der angrenzenden Gebiete Süddeutschlands. Aufl. 2. Teil II. — Tübingen 1900.
Hansgirg, A., Biologische Fragmente. —■ Bot. Centralblatt, 56. 1893.
Hegi, G., Illustrierte Flora von Mittel-Europa, V:2. — München 1926.
Håkansson, A., Studien über die Entwicklungsgeschichte der Umbellife- ren. — Lunds Univ. Årsskrift, N.F., Avd. 2, Nr 7. 1923. (Också diss., Lund 1923.)
Karsch, Flora der Provinz Westfalen und der angrenzenden Gebiete. Achte Aufl. bearb. von H. Brockhausen. — Münster (Westf.) 1911.
Karsten, H., Deutsche Flora. Flora von Deutschland, Oesterreich und der Schweiz etc. Teil II. — Gera-Untermhaus 1895.
Kronfeld, M., Ueber Anthokyanblüten von Daucus Carota. — Bot. Cen tralblatt, 49. 1892.
Kullenberg, B., Blommor och insekter. — Svensk Naturvetenskap 1956. Statens naturv. forskningsråds årsbok, 10. Stockholm 1957.
Küster, E., Zur pathologischen Morphologie der Blüten und Blütenstände. 1. Beitrag. I. Die Mohrenblüten von Daucus carota. — Zeitschrift
f. Pflanzenkrankheiten (Pflanzenpathologie) und Pflanzenschutz, 52. 1942.
Lagerberg, T., Vilda växter i Norden. Bd 3. Uppl. 2. — Stockholm 1948.
Lang, A., The effect of gibberellin upon flower formation. —- Proc. Nat. Acad, of Sei. of U.S.A., 43. 1957.
Lange, J., Haandbog i den danske Flora. — Kjobenhavn 1851. (4. Udgave 1886-88.)
Lindman, C. A. M., Bilder ur Nordens flora, III. — Stockholm 1905. —»—, Svensk fanerogamflora. — Stockholm 1918.
Molliard, M., Sur les phénoménes tératologiques survenant dans l’appareil floral de la Carotte å la suite de traumatismes. — C. R. hebd. d. séances de l’Acad. d. Sei., 172. Paris 1921.
Muschler, R., A manual flora of Egypt, 2. — Berlin 1912.
Möbius, M., Beiträge zur Biologie und Anatomie der Blüten. —■ 44. Ber. d. Senckenbergischen Naturforsch. Ges. in Frankfurt am Main. 1913.
Nelson, E., Gesetzmässigkeiten der Gestaltwandlung im Blütenbereich. Ihre Bedeutung für das Problem der Evolution. — Chermex- Montreux 1954.
Neuman, L. M., Sveriges flora (fanerogamerna). — Lund 1901.
Nicolaisen, N., Möhre, Daucus Carota sat. L. (In Roemer, T., & Rudolf,
W., Handbuch der Pflanzenzüchtung, 5.) — Berlin 1950.
Nilsson, A., Anteckningar om Landskrona-traktens flora. II. — Bot. Notiser 1952.
Nordhagen, R., Norsk flora med kort omtale av innforte treslag, pryd-og nytteplanter. — Oslo 1940.
[Patch, Edith M.], Chimera or accident of development? — Journal of
Heredity, 21. 1930.
Penzig, O., Pflanzen-Teratologie systematisch geordnet. Bd. 2. Aufl. 2. — Berlin 1921.
Pichenor, Mathilde, Modifications apportées par le greffage dans le développement du Petunia violacea Lindl. — C. R. hebd. d. séances de l’Acad. d. Sei., 244. Paris 1957.
Rafn, C. G., Danmarks og Holsteens Flora. Del 2. — Kjobenhavn 1800.
Robbins, W. W., Floral development in Daucus carota. — American Journ. of Bot., 18. 1931.
Robinson, B. L., An unusual Daucus Carota. — Rhodora, 21. 1919. Ross, H., Die Pflanzengallen (Cecidien) Mittel- und Nordeuropas, ihre
Erreger und Biologie und Bestimmungstabellen. — Jena 1911.
Russell, W., Les fleurs centrales de Daucus Carota et leurs anomalies. —- Revue générale de bot., 38. 1926.
Rohling, J. C., Deutschlands Flora ... bearbeitet von F. C. Mertensund
\V. D. J. Koch. Teil II. — Frankfurt am Main 1826.
Schinz, H., & Keller, R., Flora der Schweiz etc. Aufl. 4. — Zürich 1923.
Schröder, H., Untersuchungen über die Beeinflussung des Blütenfarben musters von Petunia hybrida grandiflora hört. — Jahrb. f. wiss. Bot., 79. 1934.
Schube, T., Flora von Schlesien preussischen und österreichischen Anteils. — Breslau 1904.
von Schwerin, F., Mitteilungen in Yerhandl. d. Bot. Yer. d. Prov. Bran denburg, 64. 1922.
Smith, E., The English Flora, 2. — London 1824.
Troll, W., Praktische Einführung in die Pflanzenmorphologie. Teil 2. Jena 1957.
Vollman, F., Flora von Bayern. — Stuttgart 1914.
Warming, E., Smaa biologiske og morfologiske Bidrag. 3. Om Skaerm- planternes Skaerm. — Bot. Tidskrift. Tredie Riekke. 1. — Koben- havn 1876-77.
Wenderoth, G. W. F., Flora Hassiaca oder systematisches Verzeichnis alle bis ietz in Kurhessen ... beobachteten Pflanzen, ... — Cassel 1846.
Willkomm, M., & Lange, Joanne, Prodromus florae Hispanicae, 3. - Stuttgartiae 1880.
SMÄRRE UPPSATSER OCH MEDDELANDEN.
Föreningens medlemmar uppmanas att till denna avdelning insända meddelanden om märkliga växtfynd o. d.
Linné-jubileet i Uppsala.
I samband med det akademiska läsårets avslutning högtidlighölls vid Uppsala universitet den 28-31 maj 1957 tvåhundrafemtioårsminnet av
Carl von Linnés födelse. De två första dagarna av jubileet ägnades åt
ett internationellt symposium över ämnet »Systematics of to-day», omfat tande korta föredrag och diskussioner över aktuella systematiska problem. President för detta symposium var prof. J. A. Nannfeldt, vice-president prof. S. Hörstadius, och som sekreterare fungerade undertecknad. Till symposiet hade inbjudits ett antal framstående utländska botanister och zoologer, av vilka närvoro: J. Braun-Blanquet (Frankrike), J. Clau
sen (U.S.A.), F. Ehrendorfer (Österrike), J. Feldmann (Frankrike),
G. Heberer (Västtyskland), R. Heim (Frankrike), J. Heslop-Harrison
(Storbritannien), J. Huxley (Storbritannien), J. Iversen (Danmark),
Z. Lorkovic (Jugoslavien), W. Ludwig (Västtyskland), Irene Manton
(Storbritannien), E. Mayr (U.S.A.), H. Merxmüller (Västtyskland),
R. Nordhagen (Norge), R. Pichi-Sermolli (Italien), B. Rensch (Väst
tyskland), R. C. Rollins (U.S.A.), F. A. Schilder (Östtyskland), C. G.
G. J. van Steenis (Holland), R. Tuomikoski (Finland), W. B. Turrill
(Storbritannien) och F. Widder (Österrike). Som representant för Hol
ländska Vetenskapsakademien kom dr L. D. Brongersma, medan dr W. B.
Turrillrepresenterade The Linnean Society of London. Dessutom deltog
ett 50-tal svenska biologer.
Symposiet öppnades på morgonen den 28 maj i det nyrenoverade Linné- orangeriet av Uppsala universitets rektor, prof. T. Segerstedt, som i samband därmed till prof. S. Ekman överlämnade en Elliot-medalj från
National Academy of Science i U.S.A. Rektor fick sedan för universitetets räkning av dr Turrill mottaga två praktfulla adresser, den ena från
Linnean Society of London, den andra från Kew Gardens. Från Svenska Läkarsällskapet överlämnade så dr F. Wahlgren Linné’s örtabok. Slut
ligen höll symposiets president sin »Presidential address».
Efter en rundvandring i Linné-trädgården och en kaffepaus på »Alma» började så den första sessionen på universitetets lärosal X. Den ägnades i huvudsak åt högre systematiska kategorier och omfattade sju föredrag.
Mayr talade om »The evolutionary significance of the systematic cate
gories»; Huxley om »Evolutionary processes and taxonomy, with special
reference to grades»; Heberer »Zum Problem der additiven Typogenese»;
Heim om »Les critéres d’ordre chimique dans 1’étude des affinités chez les
Macromycétes»; Feldmann framförde »Remarques sur la systématique
actuelle des Algues»; Tuomikoski talade »Über den heutigen Stand der
Laubmoossystematik» och Pichi-Sermolli behandlade »The higher taxa
of the Pteridophyta and their classification». Efter föredragen följde en gemensam diskussion.
Efter lunchen vidtog så session 2, som i första hand behandlade art begreppet. Den inleddes med ett föredrag av Rensch om »Die ideale Art
beschreibung»; sedan talade Manton om »The concept of the aggregate
species»; Rollinsföredrog »Some considerations of the comparative syste-
matics of vascular plant species»; van Steenis talade om »Specific delimi
tation, with special reference to tropical plants» och Ludwig om »Art
begriff und Evolution der Art unter dem Aspekt E. Geoffroys (Geoffroy-
ismus)». Efter ett kort meddelande av Erdtman, »On terminology in pollen and spore morphology», vidtog en allmän diskussion.
Symposiets andra dag började med session 3, som till större delen av handlade variation och systematiska enheter inom arten, och som omfattade ej mindre än åtta föredrag. Clausen inledde med ett föredrag om »The
function and evolution of ecotypes, ecospecies, and other natural entities»;
Turrill talade om »Genetic variation within taxonomic species»; Heslop- Harrison om »Ecological variation and ethological isolation»; Lorkovic
om »Die Merkmale der unvollendeten Speziationsstufe und die Frage der Einführung der Kategorie der Semispezies in die Systematik»; Schilder
om »Versuch einer genetischen Klassifikation der Varianten bei Bänder schnecken (Cepaea)»; Ehrendorfer om »Die geographische und ökolo
gische Entfaltung des europäisch-alpinen Polyploidenkomplexes Galium anisophyllum Vill. seit Beginn des Quartärs»; Lövkvist om »Polyploidy
and ecological differentiation» och Hedbergom »The taxonomic treatment
of vicarious taxa». Efter den gemensamma diskussionen togs så en välför tjänt lunchpaus.
Den fjärde och sista sessionen omfattade sju föredrag. Widder talade
om »Die geographisch-morphologische Methode als abgestufter Verwandt schaftstest»; Merxmüller om »Einige Probleme der Sippengliederung und
Arealbildung in den Alpen»; Iversen om »The bearing of the glacial and
interglacial epochs on the formation and extinction of plant taxa»; Du
Rietzom »The hybrid concept»; Petersenom »Pieris napi and Pieris (napi)
bryoniae, two ’Siamese twin species’»; Svärdson om »Interspecific hybrid
populations in Coregonus» och Pejler om »Taxonomic studies on plank
tonic Rotatoria». Sedan den sista sessionens ämnen debatterats, avslutades så det egentliga symposiet samma kväll med en middag på Västgöta nation. Men alla de utländska gästerna och många av de svenska deltagarna stan nade kvar under resten av jubileet.
Torsdagen den 30 maj (Kristi Himmelsfärdsdag) ordnades för de kvar varande symposiedeltagarna under ledning av prof. G. E. Du Rietz en
exkursion i Linnés fotspår. Efter bussfärd till Hågadalen mellan St. Djur gården och Fäbodarna följde man en del av Linnés »Herbatio gottsundensis» förbi Predikstolen fram till Lurbo bro med en avstickare från Predikstolen
JBei
mm
Fig. 1. Professorerna R. Nordhagenoch H. Osvald utanför Vårdsätra friluftsgård. — Foto E. Hultén30.5. 1957.
genom en granskogsdalgång upp till en hällmarkstallskog och ett orört kärr i denna. Deltagarna fick därunder tillfälle att studera försommar aspekten i Hågadalens ängar och inte minst i lunden nedanför Predikstolen. Från Lurbo fortsatte man med buss till studentkårens friluftsgård vid Vårdsätra, där lunchen väntade. Så följde ett kort besök i Vårdsätra natur park med demonstration av den blandade lövträdslunden och alskogen vid Ekolns strand, och sedan fortsatte färden med buss till Linnés Hammarby. Efter att ha gått husesyn i corps-de-logiet och besett parken samlades exkursionsdeltagarna utanför Linnés museibyggnad för att deltaga i ett sammanträde med K. Vetenskapssocieteten. Dess preses, förre överbiblio tekarien A. Grape, höll ett välkomstanförande på franska, och sedan höll prof. N. von Hofsten på engelska ett föredrag om Linnés naturupp
fattning.
De osäkra framtidsutsikterna för de vackra lundarna vid Fäbodarna och Gottsunda hade gjort ett djupt intryck på de utländska vetenskapsmännen. På förslag av dr Ramsbottom antogo de därför med acklamation en reso
lution, som i svensk översättning lyder: »Undertecknade, vetenskapsmän från många länder, samlade i Uppsala för att fira 250-årsminnet av Linnés födelse, äro efter att ha vandrat längs en av Linnés herbationes synnerligen fängslade av landskapets skönhet och dess biologiska intresse. Med tanke på dess värde ur vetenskaplig, historisk och internationell synvinkel äro
.
.
Fig. 2. Prot. R. Heim och dr J. Ramsbottom i Vårdsätra naturpark. — Foto
E. Hultén 30.5. 1957.
vi angelägna att framhålla att detta område för framtiden bör bevaras som ett naturreservat.»
Efter ett kåseri av fil. dr A. Hj. Uggla om Hammarby och om hur
Linnés liv där tedde sig anträddes återfärden till Uppsala. Här samlades man så småningom på Linnés gamla nation, Smålands, som hyllade minnet av sin vittberömde inspektor genom att — med hjälp av den församlade botaniska sakkunskapen — plantera ett vårdträd på nationens gård. Efter ett föredrag av dr Uggla om universitetets och nationernas uppkomst
samt om Linné som inspektor och efter en välkomsthälsning av förste kurator vidtog middag i nationens högtidssal. Dagen avslutades med del tagande i den traditionella kransbindningsfesten i Linné-orangeriet.
Nästa dag, fredagen den 31 maj, kom så Linné-jubileets kulmination, doktorspromotionen i närvaro av H. M. Konungen. Promotionsföreläsning hölls av prof. S. Hörstadius, som talade om »Linné, djuren och människan». I anledning av Linné-jubileet kreerades ett ovanligt stort antal natur vetenskapliga hedersdoktorer. Inom den nya matematisk-naturvetenskap- liga fakulteten promoverades en rad utländska forskare till filosofie heders doktorer, nämligen H. H. Allan (Nya Zeeland; absens), J. Braun-Blan-
quet, R. Heim, J. Huxley, Johs. Iversen, H. Lindberg, E. Mayr, R. Nordhagen, J. Ramsbottom, B. Rensch och Johanna Westerdijk
(Holland; absens). Prof. Lindberg var en av de få av de närvarande som
varit med redan vid 1907 års Linné-jubileum — av samma års promovendi
c- / \
.. ''
\
.A,
-A
Fig. 3. Prof. G. E. Du Rietz och dr J. Braun-Blanquet på Linnés Hammarby. — Foto Uppsala-Bild 30.5. 1957.
hälsades nu bl. a. de tre biologiska jubeldoktorerna professorerna N. von
Hofsten och C. Skottsbergsamt förre byråchefen K. A. Andersson. Till agronomie hedersdoktorer promoverades dr J. Clausen, prof. H. Kihara
(Japan; absens) och prof. J. L. Lush (U.S.A.; abseits). Det blev även ett antal svenska hedersdoktorer; sålunda promoverades till agronomie hedersdoktor prof. G. Turesson (absens), till medicine hedersdoktorer de
båda Linné-forskarna med. lic. T. Fredbärj och fil. dr A. Hj. Uggla. Till filosofie hedersdoktorer promoverades inom humanistiska fakulteten bl. a. Växjö-biskopen och Linné-forskaren E. Malmeström och inom matema-
tisk-naturvetenskapliga fakulteten med. lic. H. Fries.
Efter promotionen följde studentkårens traditionella uppvaktning för jubeldoktorer och hedersdoktorer, och sedan var det dags för linneanerna att bege sig till ett litet garden party i universitetets botaniska trädgård. De kvarvarande symposiedeltagarna fick här ännu ett tillfälle att utbyta
Fig. 4. Prof. R. Pichi-Sermolli och fru Maj Runepå Linnés Hammarby. — Foto E. Hultén 30.5. 1957.
åsikter och erfarenheter; allmän enighet tycktes råda om att de två sym- posiedagarna varit mycket givande.
Yid Linné-jubileets festliga final, promotionsbanketten i rikssalen på Uppsala slott, blommade åter den akademiska vältaligheten. Efter rektors välkomsthälsning fick man höra prof. Å. Malmström tala till jubeldokto
rerna, vilkas svar lämnades av förre rector magnificus prof. N. von Hor
sten. Till hedersdoktorerna talade lärt och fyrspråkigt prof. E. Ask- Upmark, och hans tal återgäldades för svenskarnas del av prof. E. Wel-
lander och för utlänningarna av prof. J. Huxley. Till de nypromove- rade talade prof. G. Gunnarsson, och deras svarstal kom genom docent
A. Önnerfors. Talet till kvinnan hölls av med. dr L. Garby, och slutligen tackade hans excellens riksmarskalken och nypromoverade jubeldoktorn
Birger Ekebergför maten. Efter kaffet dansade man så tills det ljusnade
— och så var jubileet slut. Linnés minne var firat, och betydelsen av hans
()11 insatser inom de biologiska vetenskaperna hade ännu en gång understrukits. —• Linné-symposiets föredrag och diskussionen kommer att publiceras i Uppsala universitets årsskrift.
Olov Hedberg.
On the Risks of Overstressing the Denominations of Fossil Spores as Illustrated by a Paper on the Former Distribution of
Lophosoria (“Cyathea annulata”).
In 1947 Dr. Isabel Cookson described some fossil spores, referred to
as “ Cyatheacidites annulata Cookson”, from Tertiary lignites of the Ker guelen Archipelago. In a subsequent paper (Cookson 1956) occurrences of
spores of the same kind, now referred to as “Cyathea annulata Cookson”,
in Lower Tertiary, Oligocene, and Pliocene ligneous or carbonaceous clays in Australia (Queensland, New South Wales, and Victoria) are reported. These spores are said to have a thick “exospore”, pitted and covered with irregular protuberances, especially marked on the facets of the wall of the proximal part of the spore, where they coalesce to form three conspicuously thickened areas. To this Dr. Cooksonadds the following comments: “When
first described the close agreement in every detail between C. annulata and the spores of Cyathea vestita Mart, from tropical South America was noted.
Knox(1939, p. 454) has shown that the spores of Lophosoria quadripinnata
have the same general type of construction, but lack the conspicuous ex crescences present on the facets of the proximal surface of this species. Since localised proximal thickenings are a conspicuous feature of C. annulata, there thus seems little doubt that a species of Cyathea closely related to C. vestita was relatively widely distributed in the southern hemisphere during the Tertiary.”
The above remarks may be commented on as follows. The spores in Cyathea vestita are, as far as I have been able to discern (cf. Erdtman1957,
Fig. 96, p. 56), distinctly different from the more or less unique type of spores in Lophosoria quadripinnata (cf. op. cit., Figs. 133-134, pp. 71-72). As to Mrs. Knox’s figures, the following should be observed: Figs. 75 a
and b in her paper without doubt show spores of Lophosoria quadripinnata (not Cyathea vestita, as given in the legend), whereas Figs. 74 a and b, in conformity with Mrs. Knox’s statement, illustrate spores of the same,
rather polymorphous species.1
The stratification of the fossilized part of the spore wall is not clearly shown in Cookson 1956, PI. IX and Figs. 4—5. This urges the desirability of supplementing photomicrographs with sketches showing the details of sporoderm stratification. The term “exosporium” employed by Dr.
Cookson may perhaps be misleading since in her paper no indication is
supplied to show that the outermost part of the spore wall in the alleged
1 Professor Carl Skottsbergkindly informs me that in the rain forest of Western Patagonia the habit of Lophosoria quadripinnata is appreciably different from its habit in the Juan Fernandez Islands (cf. Skottsberg in Vet. Akad. Handl. 56: 5,
Taf. 5, and in Nat. Hist. J. Fern. II, PI. 115: 1).
Cyalhea annulata is of a non-perinous nature. In undetermined cases of this nature the wail may, in a less committal way, be referred to as scleri- nous (“sclerine” comprising perine and/or exine). Spores of the same type as in Lophosoria quadripinnata have not, so far, been found in any other genus. It would, therefore, probably be possible to refer Cyatheacidites annulata Cooks, (syn. Cyathea annulata Cooks.) to Lophosoria cf. quadri pinnata (Gmel.) C. Chr.
Lophosoria is an isolated relict genus now usually not retained in the Cyatheaceae but referred to a special family, the Protocyatheaceae. The interesting findings of Dr. Cookson substantiate Professor Skottsberg’s
idea of Lophosoria being a tricentric Antarcto-tertiary element (Skotts-
berg 1956 and letter Nov. 8, 1957).
In the same paper Dr. Cooksonhas exchanged the generic name “Dacry-
diumites Cookson” for “Dacrydium”, as the pollen grains described as Dacrydiumites are closely related to those of the living genus Dacrydium. In its present compass, however, Dacrydium is heterogeneous (cf., e.g.,
Erdtman 1957) and will probably be split in the near future. Therefore,
it would probably have been better to postpone the transfer of Dacry diumites to Dacrydium.
“Amylotheca pliocenica Cookson n.sp.” is another species which can
hardly be accepted by palaeobotanists until proof has been furnished that pollen grains of the same type as those in Amylotheca do not occur in other genera. Dr. Cookson has not provided such proof.
“Alangium javanicoides Cookson” is still another species. Here points of view similar to those quoted above under Dacrydium can be advanced.
On pp. 218-19 in “An Introduction to Pollen Analysis” (Erdtman 1943,
new printing 1954) I have stressed that “the naming of fossil pollen grains and spores must be done carefully to avoid the impression of overstressing the denomination”. This is equally true today. In the same book (p. 53) it is also stressed that “the illustrations are meant to convey suggestions as to the identification of pollen grains and spores and are not meant to take the place of pollen and spore preparations”. The same holds good for all books and papers of the same kind, including the paper by Knoxwhich
was published comparatively long ago (1938) and, incidentally, contains several illustrations of somewhat doubtful value.1
REFERENCES.
Cookson, I. C., 1947: Plant microfossils from the lignites of Kerguelen Archipelago. — Rep. B.A.N.Z.A.R. Exp., Ser. A, Vol. 2, Part 8, pp. 127-142.
—»—, 1956: On some Australian Tertiary spores and pollen grains that extend the geological and geographical distribution of living genera. — Proc. Roy. Soc. Viet., Vol. 69, pp. 41-53.
1 Thus Fig. 52, p. 451 in Knox’spaper of 1938, is not Hymenophyllum tunbridgense,
which has trilete, thin-walled spores (as shown, e.g., by Mrs. Knoxherself in a later,
beautifully illustrated paper, viz. Knox1951, Fig. 5, Pi. XX), but likely a stray spore of Polypodium (?), which accidentally came along with the specimen investigated.