Skaftslamkrypa (Elatine hexandra) i Bolmen,
utbredning, tillväxt, reproduktion och styrande
faktorer
Författare: Anna Edlund
Examensarbete i biologi
Nivå: Filosofie magister examen
Nr:
Examensarbeten gjorda vid Högskolan i Kalmar, Naturvetenskapliga institutionen, och lista över dessa kan beställas via;
www.hik.se/student eller: Högskolan i Kalmar Naturvetenskapliga institutionen 391 82 KALMAR Tel 0480-44 62 00 Fax 0480-44 73 05 e-post: info@nv. hik.se
Examensarbeten gjorda på Högskolan i Kalmar finns på: www.hik.se/student
Detta är ett examensarbete och studenten svarar själv för de framlagda resultaten och slutsatserna i rapporten.
Abstract
The plants in the genus Elatine are characteristic for oligo and mesotrophic lakes. This study focuses on Elatine
hexandra one of the four species in Sweden. It is stated as vulnerable by the National Environment Agency and a
conservation plan with guidelines to preserve the plant in Sweden is under preparation. There have been a few previous studies investigating this plant in other countries but this is the first one for Sweden. The aim of this study was to investigate the distribution, growth and reproduction of E. hexandra in the lake Bolmen, and to investigate what the factors that could influence them. Factors that where included in this study were water depth, wave exposure, amount of epiphytes and sediment characteristics.
An analysis of emerging seedlings from soil seed banks revealed a maximum density of 2200 seeds /m2 to 10 cm depth. A linear regression showed that the size of the seed bank was negatively related to wave exposure (F1,3=
36,71 P=0,009, R2=0,96).
In August, the depth distribution of the plants was described in 5 transects at 7 sites. All pla nts were found to grow submerged, the depth of the plants ranged between 3-74 cm. The mean depth of the plants was
significantly more shallow then expected from an random distribution in 2 of the 7 sites (F6,55= 10,47, P<0,05,
ANOVA, Tukey, P<0,05). Further the mean depth differed between the localities and it was positively related to the water content of the surface sediment in shallow water (0-15 cm, linear regression, F1, 5=14, 43, P=0, 02, R2 = 0, 88), but not with water content of the surface sediment in deep water (15-30 cm). Mean plant depth was not correlated with wave exposure or epiphytic biomass suggesting that local sediment factors were more important in this lake.
Furthermore, a comparison with 7 randomly located sites without Elatine showed the water content of the surface sediment was higher in sites without Elatine. This supports the supposition that soft sediments are poor growth habitats for this species.
In October, when the water level had dropped to its annual minimum, plant size and seed production were examined in emerged and submersed plants at 5 sites. Plant length was smaller in emerged (25.1±7.6mm, mean±95%conf.int) than in submerged positions (40.3±8.3mm, F1, 40=14.9, P<0, 05, ANOVA). Similarly, the
number of root per node was s maller in emerged (2.2±0.2) than in submerged positions (2.8±0.2, F1, 40=14.9, P<0, 05, ANOVA). No consistent difference was found with respect to seed production per plant, 34.4±22.5 and 61.4±24.2 seeds per plant in emerged and submerged positions, respectively (Depth: F1,40=1.8, P>0.05,
ANOVA). In this case, the effect of depth was dependent on site (Depth X Site: F4, 40= 2.7, P<0, 05, ANOVA).
Annual and interannual variations in water regime in relation to distribution and size of the seed bank are proposed to be of key importance for conservation of E. hexandra . Further studies on the ability to germinate above water in Sweden may shed light on the finding that no plant occurred above water level in the present study. An investigation of the seed banks in former habitats is proposed to answer if the species have abandoned the habitat or if the habitat no longer is suitable while there are still seeds resting in the seed bank.
Innehållsförteckning
INTRODUKTION ... 5
MATERIAL OCH METODER ... 8
SJÖBESKRIVNING...8
EXPONERINGSBERÄKNING...9
VATTENNIVÅDATA...10
FRÖBANKSUNDERSÖKNING...10
DJUPUTBREDNING OCH TÄTHET...12
JÄMFÖRELSE MED LOKALER UTAN SKAFTSLAMKRYP A...14
VATTENNIVÅNS PÅVERKAN PÅ TILLVÄXT OCH REP RODUKTION...14
RESULTAT ...16
FRÖBANKSUNDERSÖKNING...16
DJUPUTBREDNING OCH TÄTHET...17
JÄMFÖRELSE MED LOKALER UTAN SKAFTSLAMKRYP A...21
VATTENNIVÅNS PÅVERKAN PÅ TILLVÄXT OCH REP RODUKTION...21
VATTENNIVÅFÖRHÅLLANDE I BOLMEN...23
DISKUSSION...24
FRÖBANKSUNDERSÖKNING...24
DJUPUTBREDNING OCH TÄTHET...25
JÄMFÖRELSE MED LOKALER UTAN SKAFTSLAMKRYP A...27
VATTENNIVÅNS PÅVERKAN PÅ TILLVÄXT OCH REPRODUKTION...27
VATTENNIVÅFÖRHÅLLANDE I BOLMEN...28
SLUTSATSER OCH IDÉER FÖR BEVARANDE AV SKAFTSLAMKRYPA...29
REFERENSER ...31
BILAGA 1) KARTOR ...33
BILAGA 2) FOTON...35
Introduktion
Oligo- mesotrofa sjöar (Natura 2000 naturtyp 3130) är en av de miljöer som innefattas av art habitat direktivet inom EU. Dessa sjöar kan vara extra känsliga för försurning då de naturligt redan har ett lågt pH och en svag buffringsförmåga (Naturvårdsverket 2003). I oligotrofa sjöar finns också de växtarter som kanske har svårast att klara de konsekvenser som övergödning innebär som till exempel skuggning från utbredande vassbestånd och ökad grumlighet i vattnet.
För denna naturtyp är arterna inom släktet slamkrypor (Elatine) typiska karaktärsarter (Naturvårdsverket 2003). Släktet har fyra arter i Sverige; slamkrypa (E. hydropiper), tretalig slamkrypa (E. triandra), nord slamkrypa (E. orthosperma), och skaftslamkrypa (E. hexandra). Denna studie kommer att behandla arten skaftslamkrypa som, liksom de övriga arterna i Sverige, växer på bottnar eller stränder med tunnare gyttje lager och är en mycket oansenlig vattenväxt. Plantorna blir inte mer än några cm långa, i undantagsfall upp till 15 cm (figur 1). Skaftslamkrypa klarar av att blomma och frösätta både på land och i vatten (Salisbury 1967). Blommorna som växer från bladnoderna är skaftade och med sex ståndare, vilket skiljer den från de övriga slamkryporna. Blommorna är tillbakabildade med små kronblad, de kan fortfarande korspollineras men huvudsakligen är de självpollinerande, speciellt under vatten då de genom denna strategi inte är i behov av att blommorna kommer upp till ytan för pollinering (Sculthorpe 1985)(bilaga 2 figur 3b). Mängden frön per kapsel från vattenlevande plantor är oftast färre då dessa huvudsakligen kommer från självpollinerade blommor (Salisbury 1967).
Figur 1. Skiss av skaftslamkrypa (Ill. A. Edlund 2008)
Skaftslamkrypa har uppmärksammats som en hotad art. Artdatabanken har kategoriserat den som sårbar på svenska rödlistan då dess förekomst gradvis reducerats, både geografiskt och i
populationsstorlek (Gärdenfors 2005). Den kan hittas på västkusten men har även rapporterats in i landet, mot Mälardalsområdet och upp mot Värmland (Artdatabanken 2007). Den förekommer sällsynt i västra och sydvästra Norge samt på Jylland i Danmark (Thyssen 1990, Artdatabanken 2007).
Länsstyrelsen i Kronoberg län, där skaftslamkrypa också växer, har fått i uppdrag av Naturvårdsverket att sätta upp ett åtgärdsprogram för arten. Sjön Bolmen i Kronobergs län är den sjö med flest observationer av skaftslamkrypa . Här hittades den på över 50 lokaler kring sjöns sydvästra och östra del under en inventering av klockgentiana (Gentiana
pneumonanthe) under 2006 (Wahlström K. opubl). Detta är anledningen till att sjön valdes för
denna studie.
Det har gjorts få undersökningar av skaftslamkrypa och resterande arter av släktet Elatine i Europa. En studie om skaftslamkrypans reproduktion och biologi gjordes i England på 1960-talet (Salisbury 1967). Gronings och odlingsförsök visade att plantor som vuxit barlagt även producerade fler frön samt att ljusintensitet var viktig för groningsframgången. I Nederländerna undersöktes utbredning av Elatine och dess miljöer för att se om möjligheten fanns för att kunna använda dem som indikatorer för olika klimat och habitat under arkeologiska studie r (Brinkkemper m.fl. 2008). Den har ökat i Nederländerna under 1900-talet, men för få subfossila fynd av den finns för att den ska kunna användas som klimatindikator i arkeologiska sammanhang.
Bernhardt m.fl. (2008) granskade två olika metoder för fröbanksundersökning av arter som växte i fiskdammar i Österrike. En av arterna som undersöktes var just skaftslamkrypa. Detta försök hade två syften, dels att avgöra vilken av metoderna groning och direkt räkning av frön som bäst beskrev fröbanken, men också att genom inventering konstatera vilka växtzoner arterna förekom i. Den fröbanksmetod som de fann var bäst var att direkt räkna antalet frön i fröbanken, en metod som kan vara tidskrävande speciellt med frön av liten storlek.
I Sverige har skaftslamkrypa inventerats, men ingen stud ie har genomförts som beskriver utbredning och reproduktion eller vilka faktorer som kan tänkas styra dessa.
Vissa kritiska händelser under plantornas livscykel som till exempel fotosyntetiserande förmåga, groning, reproduktion och överlevnad under den minst fördelaktiga årstiden är styrande i alla typer av miljöer (Hutchinson 1975). En undersökning av skaftslamkrypans
olika livsfaser och kopplingen till de faktorer som kan påverka dessa kan ge en ökad förståelse för dess krav på habitat vilket är viktigt för bevarande av arten.
Den faktor som kan ha störst påverkan på vattenväxter som skaftslamkrypa är exponering av lokalen för de vågor som orsakas av vinden. Denna störningsfaktor är både direkt i form av frötransport och skador på plantor, men även indirekt i form av erosion, transport och avsättning av sediment, näringsämne n och organiskt material (Keddy 1982). En ökad exponering kan också vara positiv då bortsköljning av finare sediment och detritus ökar som annars skulle kunna begrava skaftslamkrypa (Brinkkemper 2006). Sedimentkaraktärer som mängden organiskt material, vattenhalt och kornstorlek är följaktligen sammankopplade med exponeringen men även med varandra då finare kornstorlekar bidrar till större möjligheter för sedimentet att hålla kvar vatten. Finns de finare kornstorlekarna kvar är också chansen stor att det organiska materialet även finns kvar , vilket bidrar till högre näringshalt i sedimentet. Rörelse i vattnet orsakat av vågor kan även innebära att mängden påväxt minskar på växter (Weisner m.fl. 1997). Påväxt kan tänkas vara en negativ stressfaktor för skaftslamkrypa då påväxterna konkurrerar om ljus med den växt de sitter på (Koch 2001). Även vattendjup kan indirekt innebära stress för plantorna, då vattnet begränsar ljustillgången och följaktligen möjligheterna för fotosyntes. Ljustillgången kan även minska då högre grad av exponering rör upp sediment från botten (Madsen m.fl. 2001). Som respons på en minskad ljustillgång kan plantornas kompensera med en större yta (Hutchinson 1975). Hos systerarten tretalig slamkrypa (Elatine triandra) ökade längden på internoderna och även bladen blev större hos vattenlevande exemplar (Sculthorpe 1985). För att klara av att förankra sig i det mer rörliga bottensedimentet kan rötterna bli längre och mer förgrenade under vatten, vilket skulle bidra till en högre rot/skott kvot hos plantor som växer under vatten i förhållande till de som växer barlagt (Sculthorpe 1985)
Syftet med denna studie är att undersöka utbredning och reproduktion hos skaftslamkrypa i Bolmen och hur nämnda faktorer kan tänkas påverka. Studien har delats in i följande delstudier:
1) Fröbanksundersökning
Hur stor är fröbanken av skaftslamkrypa på kända lokaler från 2006. Finns det någo t samband mellan den uppskattade populationstätheten och storleken på fröbankerna? Kan substratet de förekommer i och exponering påverka fröantalet i fröbanken?
2) Djuputbredning och täthet
Hur stor är abundansen av plantor på olika lokaler? På vilka djup växer skaftslamkrypa i Bolmen och hur är de fördelade inom lokalerna? Är medelväxtdjupet slumpmässigt eller påverkas detta på något sätt? Kan exponeringsgrad, sediment (glödförlust, vattenhalt, kornstorlek), samt påväxt påverka abundans och växtdjup.
3) Jämförelse med lokaler utan skaftslamkrypa (referenslokaler) i Bolmen
Finns det skillnader i exponeringsgrad och substrat (glödförlust, kornstorlek samt vattenhalt) mellan lokaler med och lokaler utan skaftslamkrypa? Kan dessa faktorer förklara närvaron av skaftslamkrypa på en lokal och därmed ge möjlighet att bestämma om alla lämpliga habitat är koloniserade.
4) Vattennivåns påverkan på tillväxt och reproduktion
Hur påverkar vattennivån plantors storlek och reproduktionsframgång? Hur mycket påverkas detta av lokalen de växer på?
5) Vattennivåförhållande i Bolmen
Vilka vattennivåer förekommer i Bolmen över året? Hur förhåller sig dessa till resultatet från fröbanksundersökningen samt undersökningen om djuputbredning? Kan man utifrån detta dra några slutsatser om skaftslamkrypans preferenser av vattennivåer? Växer de främst över eller under vatten?
Material och metoder
Sjöbeskrivning
Bolmen är belägen i hörnet mellan Kronoberg, Halland och Jönköpings län, i södra utkanterna av det småländska höglandet (figur 2). Bolmens strandlinje är 380 km lång och domineras av långsluttande blockiga stränder, men även klapperstensstränder och sandstränder förekommer (se figur 4, bilaga 2). Långsluttande stränder ger möjlighet för en längre strandzon när vattennivån sänks, vilket ger ett större område för skaftslamkrypa att etablera sig på.
Sjön har ett avrinningsområde på 1640 km2 där hälften av vattnet kommer till sjön från två inflöden i sjöns norra del. Den regleras även genom en damm vid utflödet sedan 1940-talet. De satta regleringsgränserna tillåter en variation på 1,4 m, mellan 141,9-140,5 meter över havet (m.ö.h.)(Hein 1999) . Alla vattendjup som anges i denna undersökning har standardiserats till en referensnivå vid 141,25 m.ö.h., som var medelvattennivån i Bolmen under juli-september 2008.
Bolmen skiljer sig mellan den norra och den södra. Medeldjupet är 8 m och maxdjup et 37 meter i södra delen medan det är 5-6 m respektive 13 m i den norra delen. Skillnaden i djup tillsammans med det norra inflödet bidrar till olika näringshalter, med högsta värden i den norra delen av sjön (Hein 1999).
Figur2. Översiktskarta över södra Sverige med Bolmen markerad samt uppförstorad med de lokaler som använts
under de olika undersökningarna markerade. För koordinater på dessa lokaler se bilaga 1.
Exponeringsberäkning
Vinddata från 1990-2008 användes från Torups väderstation 35 km väster om Bolmen (SMHI 2008). Exponeringen (E) beräknades enligt formeln
Exceedance är den årliga andelen vindar =5 m/s i varje 10° vinkel och fetch är avståndet till motstående strand för samma vinkel (Keddy 1982; Weisner 1987). I figur 3 redovisas exceedence i de olika vindriktningarna, där den dominerande vindriktningen är sydvästlig (figur3).
fetch
exceedence
E
i i= i° ° × =∑
10 36 1 100,00% 10,00% 20,00% 30,00% 40,00% 50,00% 360 10 20 30 4 0 50 60 70 80 90 100 110 120 130 140 150 160 170 180 190 200 210 220 230 240 250 260 270 280 290 300 310 320 330 340 350
Figur 3. Vindfrekvenser =5 m/s för Torup väderstation 1990-2008 (SMHI 2008)
Vattennivådata
Vattennivådata för Bolmen 1997-2008 hämtades från EON AB (EON opubl) som har dagliga mätningar vid Sunnaryd och Bolmen i norra respektive södra delen av sjön. För varje inventeringslokal togs ett vattennivå värde fram genom att interpolera vattennivån mellan de båda mätstationerna så att varje latitud får ett mätvärde. Varje lokals vattennivå vid inventeringstillfället kunde därefter uppskattas.
Fröbanksundersökning
Fem lokaler i Bolmen valdes baserat på en uppskattad populationstätheten under 2006. Urvalet gjordes för att få spridning bland tidigare populationstätheter (Wahlström K. opubl.). För lokalernas placering se figur 3 bilaga 1. Sedimentprover togs 28-29 april 2008. På varje lokal slumpades fem stycken kvadratmeter stora rutor ut inom vattendjupsintervallet 0-40 cm. Vid provtagningen var vattennivån 27 cm över referensnivån (141,25+0,27=141,52 m.ö.h.), vilket innebär att proverna insamlades både i den barlagda och dränkta stranden ner till 12 cm vattendjup senare under växtperioden (juli- september) . Tio delprov togs per ruta med en jordborr (ø=2 cm), ned till 10 cm sedimentdjup. Dessa delprov sammanslogs sedan till 2 prov per ruta. Ett stort antal mindre delprov ger en bättre representation av fröbanken samtidigt som en sammanslagning av proverna minskar odlingsytan (Thompson m.fl. 1997)
Vid varje lokal togs även tio slumpmässiga prov inom provtagningsintervallet. Dessa slogs samman till ett prov per lokal för bestämning av glödförlust. Proverna förvarades vid +4°C i ca 4 veckor innan de noggrant blandades till ett sammelprov. Fyra delprov per sammelprov (lokal) togs ut och analyserades. Torkning och bränning av prover följde svensk standard (SS 028113).
Fröbanksproverna togs in till laboratoriet för sållning enligt Ter Heerdt m.fl. s metod (1996). Proverna sållades först genom ett 4 mm såll för att sortera bort grövre material sedan genom ett 0,5 mm såll ner i ett 0,125 mm såll. Fraktio ner mellan 0,5 till 0,125 mm som kvarstod vaskades för att skilja lättare organiskt material från det tyngre minerogena materialet. Detta gjordes genom att de överfördes till en behållare som fylldes med vatten. Efter omrörning kunde vattnet och det lättare organiska materialet filtreras igenom ett 0,125 mm såll, där det organiska materialet blev kvar. Detta upprepades tills inget mer organiskt material följde med vattnet.
Aluminiumformar 8×10 cm breda och 5 cm höga fylldes till ca 2-3 cm med det minerogena materialet underst i formarna och det vaskade organiska materialet överst. Proverna planterades en vecka efter provtagning och placerades sedan i ett kontrollrum med ljus 24 timmar/dygn (ca 30 µmol m-2 sek-1) fotosyntetiskt aktivt ljus).Temperaturen i kontrollrummet höll ca 20-25º C och den relativa luftfuktighet var ca 30-50 %. Substratet hölls fuktigt de första tre veckorna, men vattennivån ändrades sedan till att ha konstant dränkning med 1-2 cm vatten över substratnivån. Efter 2 veckor syntes de första plantorna i krukorna. Plantorna räknades en gång i veckan från det att de börjat synas i formarna. Skaftslamkrypa kunde inte artbestämmas förrän 7 veckor in i försöket vilket ger en osäkerhet om hur många som kan ha dött under dessa första veckor. Individuella plantor som kom upp var mycket små och inte svåra att skilja från varandra. Tre andra arter tillsammans med skaftslamkrypa dominerade i burkarna. Två av dessa var rosettväxande och inte förväxlingsbara med skaftslamkrypa medan den tredje var förväxlingsbar när enbart hjärtbladen var ut vecklade (se bilaga 2 figur 3). Förväxlingsarten kunde senare artbestämmas till notblomster (Lobelia dortmanna) Därmed finns en osäkerhet om antalet skaftslamkrypa som grodde men dog innan de artbestämts. Då inga plantor beskuggade varandra tilläts de vara kvar i burkarna under hela försöket. Sedimentytan luckrades försiktigt efter 8 veckor för att ge ljus åt eventuellt begravda frön. Försöket avslutades den 20 juli efter 11 veckor.
Fröbankstätheten beräknades genom att summera antalet plantor för de två delproven från varje provruta och multiplicera med 318,31 för att uttrycka frötätheten som antal grodda . plantor/ m2 ner till tio cm sedimentdjup. Skillnader i fröbankstäthet mellan lokaler testades med ANOVA med log(x+1) transformerade värden. Tukey’s post hoc test användes för att avgöra vilka lokaler som skiljde sig från varandra. Levenes test för homogenitet i variansen visade att log transformering av data var nödvändig för att förbättra residualfördelningen.
Pearson’s Produk t-moment korrelations test användes för att testa sambanden mellan log (x+1) fröbankstäthet och faktorerna exponering (km), glödförlust (%) samt den uppskattade populationsstorleken 2006. Exponeringsfaktorn kvadratrots transformerades, glödförlusten arcsin transformerades och populationsstorleken log+1 transformerades för att normalfördela data.
Djuputbredning och täthet
Inventeringen utfördes 10-30 augusti 2008. Sju lokaler valdes utifrån de lokaler som tidigare lokaliserats kring Bolmen (Wahlström K. opubl.). Varje lokals position bestämdes med en GPS enhet. För lokalernas placering se bilaga 1 figur 1. På varje lokal markerades en strandlinje där 5 transekter placerades slumpvis i rät vinkel mot strandlinjen. För att med säkerhet separera de olika transekten placerades de med minst två meters mellanrum längs strandlinjen. Denna linje varierade från 10 till 50 meters längd beroende på strandens utformning. Längden valdes så att habitatets strandtyp blev representerad. I varje transekt eftersöktes först plantor från vattenbrynet upp till den del av stranden som markerade högvattennivån. Därefter söktes plantor från vattenbrynet ut till 60 cm vattendjup i ett 60 cm brett band utmed transekten med hjälp av en vattenkikare. Vid varje påträffad planta noterades vattendjupet. Då plantor av skaftslamkrypa kan växa tätt där enskilda plantor ej kan urskiljas räknades dessa som en planta. Växtinventeringen kompletterades (23-26/9 2008) med inmätning av bottennivåer utmed transektlängden. Vattendjupet mättes varje meter och standardiserades mot referensnivå n (se ovan). Då vattennivån sjunkit sedan augustiinventeringen extrapolerades vattendjupen utmed den barlagda sträckan mellan vattenbrynets läge i september och i augusti.
Påväxtalger är en av de faktorer som kan påverka ljustillgången och därmed hur djupt plantorna kan tillväxa. Notblomster valdes som referensväxt för undersökning av påväxt för att undvika att skada fler plantor av skaftslamkrypa än nödvändigt och för att dess blad var lättare att skrapa. Påväxt undersöktes på de sju inventerade lokalerna i Bolmen. Tre prover med två plantor per prov insamlades från 15-30 cm djupintervall i samband med inventeringen i augusti. Påväxter skrapades av bladen samma dag som provtagning. Totalt skrapades åtta blad per prov och både över och undersidan skrapades då dessa båda kan tänkas ha tillgång till ljus. Enbart blad som var nya för året valdes för skrapning. Efter avskrapning filtrerades påväxterna ner på ett glasfiberfilter med porstorleken 1.2 µm (Whatman GF/C). De förpackades ljustätt samt frystes ner för förvaring fram till
klorofyllanalysen(Jespersen och Christoffersen, 1987). Bladen sparades för inscanning och ytbestämning med programmet ImageJ version 1.41k (Rasband, W.S.). Ytan som bestämts dubblades då både över och undersida på bladen skrapats. Filtren med proverna extraherades i 6 ml 96 % etanol i 24 timmar enligt metod av Jespersen & Christoffersen (1987). Flourosensen (µg klorofyll a / ml etanol)mättes med en Turner 10AU flourometer. Halten klorofyll a beräknades genom att multiplicera med spädningsfaktorn (6 ml). Slutligen kunde halten klorofyll a per mm2 bladyta bestämmas genom att dividera varje provs totala bladyta med den totala halten klorofyll a.
Djupfördelningen kan också påverkas av sedimentets egenskaper. I samtliga sju lokaler insamlades sedimentprover med en jordborr (ø=2 cm) för bestämning av vattenhalt, GMSand (%) och glödförlust under perioden 10-30/8. Tio prover insamlades slumpmässigt från 0-15 cm samt 15-30 cm vattendjup.För lokalen Tannåker togs enbart sedimentprover i intervallet 15-30 cm. Proverna förvarades i +4°C i 8 veckor fram till provtagning. Vattenhalten bestämdes genom att proverna sedimenterades i 24 timmar inna n överflödigt vatten togs bort. Sedimentet omrördes noggrant innan 4 deglar per prov fylldes och den blöta vikten kunde bestämmas. Torkning och bränning av prover följde svensk standard (SS 028113).
GMSand (%) används i detta arbete för att beskriva torrviktsandelen av grövre kornstorlekar (2-0,25 mm) i mineraljordens sand-silt- lerfraktion (< 2 mm). För att bestämma denna användes de redan brända proverna från djupare vatten (15-30 cm). Dessa silades genom ett 2 mm såll och vägdes. Proverna silades sedan igenom ett 0,25 mm såll där återstoden i sållet vägdes och den grövre fraktionen kunde bestämmas.
För att fastställa om medeldjupet som plantorna växte på skilde sig från en slumpmässig fördelning testades plantornas medeldjup först mot bottendjupet (medeldjupet på botten för transektet). Data var insamlade för varje transekt, vilket gav 5 värden per djupvariabel att testa med en tvåvägs ANOVA där djup typ (för växt respektive lokal) var fix och lokal en slumpmässig variabel. Levenes test visade signifikanta skillnader i varianser som dock var så små att de inte bedömdes kräva transformering av data. Tukey’s post hoc test användes för att avgöra i vilka lokaler som de båda djupen var skilda från varandra. Pearson’s Produkt-moment korrelations test användes för att testa samband mellan medeldjupet och faktorerna exponering (km), glödförlust (%), vattenhalt (%),GMSand (%) samt påväxtbiomassa (mätt som µg klorofyll a per mm2 bladyta)
Tätheten i varje transekt beräknades som antal plantor på den bottenyta som utgjordes av transektlängden från 0 -60 cm djup och transektbredden 60 cm. Detta gav 5 transekt per lokal som kunde användas för analys av skillnader i täthet (antal plantor / m2) mellan lokaler. Denna skillnad testades med en envägs ANOVA på log (x+1) transformerade data. Levenes test visade signifikanta skillnader i varianser som krävde transformering. Pearson’s Produkt-moment korrelations test användes för att testa samband mellan tätheten och faktorerna exponering (km), glödförlust (%), vattenhalt (%),GMSand (%) samt påväxtbiomassa (mätt som µg klorofyll a per mm2 bladyta) . Faktorn glödförlust (15-30 cm) kvadratrots transformerades innan analys för att få bättre normalfördelning på data.
Jämförelse med lokaler utan skaftslamkrypa
De inventerade lokalerna jämfördes med slumpartade lokaler i Bolmen där skaftslamkrypa inte förekom 2008. Förutom de sju inventeringslokalerna användes ytterligare sex lokaler som tidigare strukits för att de inte uppfyllde kravet på djupintervall 0-60 cm. Dessa lokaler provtogs samtidigt och på samma sätt som de övriga inventeringslokalerna. För denna undersökning slumpades även sju referens lokaler ut i Bolmen. Då undersökningen hittills berört sydvästra samt östra delen av Bolmen begränsades referenslokalerna även till detta område. För karta över referenslokaler se figur 2 bilaga 1. Faktorerna som jämfördes var exponering, glödförlust och vattenhalt . Sedimentprover för referenslokaler där skaftslamkrypa inte förekom 2008 togs i samband med plantinsamlingen den 20 september. För denna undersökning togs enbart sedimentprover på 0-10 cm djup, vilket motsvarar standarddjupet 20-30 cm. Två av lokalerna provtogs strax ovanför vattennivån då dessa var klapperstenstränder med begränsade provtagningsmöjligheter. För dessa lokaler beräknades av denna anledning inte vattenhalten. Proverna förvarades i +4°C i två veckor före analys. Analysmetoder följer beskrivning ovan.
Test av skillnader utfördes med t-test för exponering och GMSand. Variansen testades med Levenes test. Faktorerna vattenhalt och glödförlust uppvisade båda stora avvikelser från normalfördelning och analyserades med ett ickeparametriskt Mann-Withney U-test.
Vattennivåns påverkan på tillväxt och reproduktion
I slutet av tillväxtperioden (3 oktober 2008) insamlades plantor för att jämföra storlek och reproduktionsframgång hos barlagda (nivå 141,25-141,10 m.ö.h.) och dränkta plantor (nivå 141,10-140,90 m.ö.h.)på fem lokaler i Bolmen. Medelva ttennivån hade sjunkit till 141,04
m.ö.h, ca 20 cm under referensnivån. Lokalerna som valdes var de inventerade lokalerna Odensjö Södra, Odensjö Norra, Skeda samt Hölminge. Den femte lokalen låg i anslutning till lokalen i Hölminge och hade tidigare varit aktuell som inventeringslokal men saknade möjlighet för transekt ut till 60 cm.
Ett krav vid valet av lokaler var att plantor fanns inom de två djup nivåerna. Även antalet plantor på lokalen var viktig att ta hänsyn till då den negativa effekten på populationen blir mindre om man tar bort plantor från stora populationer. På varje lokal insamlades slumpmässigt 5 plantor som växte barlagt och 5 plantor som växte dränkt, alternativt som barlagts under de föregående dagarna (max 5 dagar). Plantorna undersöktes inom tre veckor från det att de samlats in. Preparermikroskop samt mm papper användes för undersökningen. På varje planta räknades antalet rötter på fem noder med start från den äldsta noden. På en slumpvis vald rot på varje nod mättes rotlängden. Antalet noder och längden på internoderna på plantorna räknades också. Slutligen räknades antalet blommor, antalet kapslar samt antalet frön i kapslarna. Frön som var tydligt utvecklade men ännu omogna räknades in i fröantalet då de antogs kunna mogna under den resterande tiden på växtperioden (se bilaga 2 bild 3 för bild på frön).
Plantkaraktärer som testades för skillnader mellan djupen var längd (mm), antal noder, medel rotlängd per planta (mm), medel antal rötter per nod, frön per kapsel samt frön per planta. Dessa testades med en tvåvägs ANOVA med växtdjup som fix kategorisk variabel med två nivåer (dränkt resp. barlagt) och lokal som slumpmässig variabel. Levenes test visade skillnader i varians för antal frö per kapsel, frön per planta samt längd. Dessa data transformerades innan analys. Frön per kapsel samt frön per planta log y+1 transformerades då de innehöll nollvärden.
Pearson’s Produkt- moment korrelations test användes för att testa samband mellan de olika växt parametrarna och faktorerna exponering (km), glödförlust(%), vattenhalt(%),GMSand (%) samt påväxtbiomassa (mätt som µg klorofyll a per mm2 bladyta) .
Resultat
Fröbanksundersökning
Antalet vilande frön i fröbanker (fröbankstätheten) på 5 olika lokaler i Bolmen varierade mellan 0 och 2200 frön per m2 ned till 10cm djup i sedimentet. Fröbankstätheten skiljer sig mellan de olika lokalerna (F3,16= 4,56, p=0,02, ANOVA, tabell1). Lokal 1 och 5 är skilda från
varandra (P=0,02, Tukey) medan lokal 2 och 4 inte skiljer sig från någon annan lokal (P>0,05, Tukey). I lokal 3, referenslokalen utan förekomst 2006, grodde inga frön i fröbanken och lokalen inkluderades inte i variansanalysen (figur 4).
Tabell 1 Effekt av lokal på antalet vilande frön i fröbanken för 5 lokaler i Bolmen. Analysen är en ANOVA på
log x+1 transformerade data.
Variabel Effekt SS Error SS totalt SS DF Error Df totalt Df MS Error MS F P Fröbankstäthet (antal grodda frö/m2 ned till 10 cm sedimentdjup) Lokal 21,01 24,54 45,55 3 16 19 80,84 1,53 4,56 0,02 0 500 1000 1500 2000 2500 1 2 3 4 5 antal frö/m2 A B AB AB
Figur 4. Fröbankstäthet (antal vilande frö/m2 ned till 10 cm sedimentdjup) hos fem lokaler i Bolmen (1) Skällandsö, 2) Tannåker 3) Bolmstad 4) Jättabron 5) Odensjö). Felstaplar anger standardavvikelse (SD) (N= 5). Bokstäver på staplar anger gruppering (P<0.05, Tukey).
Förklaringsfaktorerna glödförlust (gf) och förekomstyta 2006 (f06) var inte korrelerade med fröbankstätheten (R2 gf = 0,09, R2 f06 = 0,84, Pearson Produkt- moment test, P>0,05).
Lokalernas exponering (EXP) är däremot negativt korrelerad med fröbankstätheten (FB). Sambandet kan uttryckas med ekvationen, FB = -2,45vEXP +6,91 (F1,3= 36,71 P=0,009,
Linjär regression, R2= 0,96)(figur 5). Det negativa sambandet är signifikant även om lokalen utan frö utesluts ur analysen.
0,0 0,5 1,0 1,5 2,0 2,5 3,0 3,5 4,0 1 1,2 1,4 1,6 1,8 2 2,2 2,4 2,6 2,8 3 exponering (km)
log+1 frötäthet (plantor/m2)
v
Figur 5. Samband mellan log+1 transformerad fröbankstäthet (log10(plantor/m2))och vexponering (km) för fem
lokaler i Bolmen (F1,3= 36,71 P=0,009, Linjär regression, R2= 0,96)
Djuputbredning och täthet
Under inventeringen av 7 lokaler i Bolmen under augusti månad lokaliserades plantor mellan 3-60 cm djup (figur 6). I Hölminge hittades dessutom 4 plantor som växte djupare än 60 cm. Dessa plantor inkluderades inte i den statistiska analysen då växterna inte systematiskt eftersöktes på större vattendjup än 60 cm i transekterna. Denna lokal och den i Tannåker var de enda med plantor i intervallet 50-60 cm (figur 6). Maxdjupet för plantorna i dessa transekter var 74 cm. Antalet plantor för varje lokal varierade från 17 (Kronobäck) till 395 (Odensjö södra).
Tabell 2. Medel för varje lokal ± 95% konfidensintervall för växtdjup och bottendjup i de sju inventerade
lokalerna i Bolmen.
Växtdjup (cm) Bottendjup (cm)
Inventeringslokal
medel ± 95% konfidensintervall medel ± 95% konfidensintervall
Odensjö södra 13,68 ± 0,45 30,53 ± 1,82 Löckna 21,45 ± 3,16 32,00 ± 3,59 Odensjö norra 14,45 ± 0,81 22,11 ± 3,11 Hölminge 35,28 ± 1,4 31,46 ± 2,47 Skeda 32,03 ± 2,32 26,65 ± 2,80 Kronobäck 34,65 ± 2,82 28,80 ± 3,58 Tannåker 28,03 ± 1,83 34,03 ± 2,40
De olika lokalerna skiljde sig åt i fråga om djuputbredning och ingen av lokalerna hade plantor som växte i alla djupintervall vilket vore fallet om fördelningen var slumpmässig. I Odensjö norra samt Kronobäck fanns mer än 50 % av alla plantor inom ett intervall mellan 10-15 resp. 35-40 cm vattendjup. I Hölminge var utbredningen jämnast fördelad (variations koefficienten (CV)=10%), inget djupintervall representerade mer än 20 % av alla plantor (figur 6).
-5 5 15 25 35 45 55 0 -5 5-10 10-15 15-20 20-25 25-30 30-35 35-40 40-45 45-50 50-55 55-60 djupintervall(cm) andel plantor (%) A B C D E F G
Figur 6. Andelen plantor (%) vid olika vattendjup för sju lokalerna (a-g) i Bolmen. Vattennivån 141,25 användes som
nollnivå, se vidare material och metoder (a) Odensjö Södra N=395, CV=18% b) Odensjö Norra N=96, CV=17% c) Löckna N=22, CV=24% d) Hölminge N=97, CV= 10% e) Tannåker N=92, CV=28% f) Skeda N=78,CV=16% g) Kronobäck N=17, CV=24%)
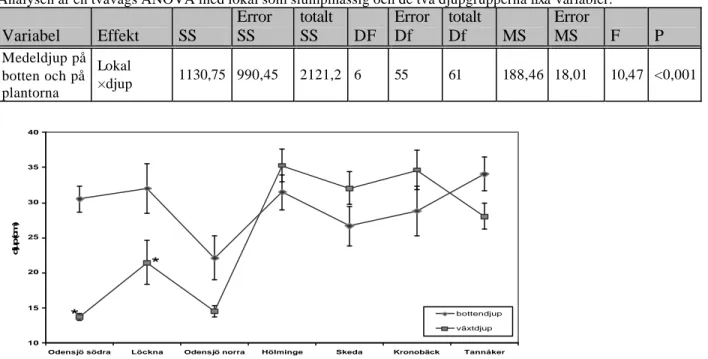
Medelväxtdjupet i varje transekt jämfördes med medelbottendjupet för att fastställa om plantornas växtdjup var slumpmässig. Plantornas medelväxtdjup skilde sig från bottnens medeldjup i två av de sju inventerade lokalerna (F6,55= 10, 47, P<0,001, ANOVA, tabell 3).
Dessa lokaler var Odensjö södra (OS) samt Löckna (L) (POS = 0,0001,PL=0,0075, Tukey).
Tabell 3. Effekten av lokal ×djup på medeldjupet på botten och på plantor av skaftslamkrypa i Bolmen.
Analysen är en tvåvägs ANOVA med lokal som slumpmässig och de två djupgrupperna fixa variabler.
Variabel Effekt SS Error SS totalt SS DF Error Df totalt Df MS Error MS F P Medeldjup på botten och på plantorna Lokal ×djup 1130,75 990,45 2121,2 6 55 61 188,46 18,01 10,47 <0,001 10 15 20 25 30 35 40
Odensjö södra Löckna Odensjö norra Hölminge Skeda Kronobäck Tannåker
djup (cm)
bottendjup växtdjup
*
*
Figur 7. Plantornas medelväxtdjup samt lokalernas medelbottendjup) i sju lokaler i Bolmen. Felstaplarna anger
95% konfidensintervall. * anger lokaler där det finns skillnader mellan medelvärden på växtdjup och bottendjup inom lokalen (P< 0,05).
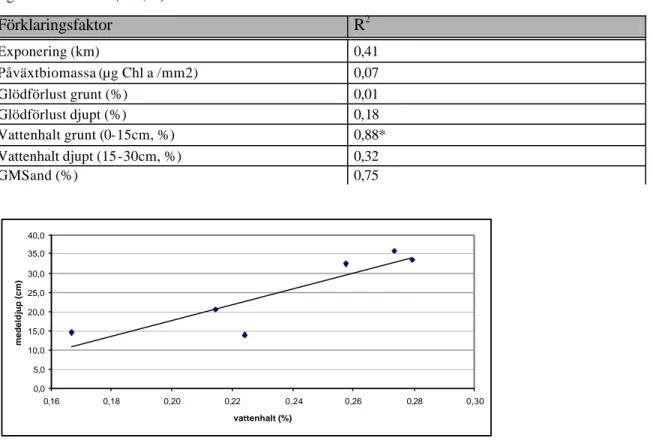
Faktorerna exponering, glödförlust, klorofyll a eller GMSand (%) var inte korrelerade med plantornas medeldjup på lokalerna (tabell 2). Sedimentets vattenhalt på grundare botten (VH)
(0-15 cm) visade ett positivt samband med plantorns me deldjup (PM). Sambandet kan uttryckas med ekvationen PM= 205,37VH - 23,407 (F1,5=14,43, P=0,02, linjär regression, R2 = 0,88, figur 8). Detta samband finns inte mellan vattenhalten på djupare botten (15-30 cm) och medeldjupet på plantorna (tabell 4).
Tabell 4. Korrelation mellan medeldjupet för plantorna i de 7 inventerade lokalerna och förklaringsvariablerna
exponering (km), påväxtbiomassa (µg chl a /mm2 bladyta), glödförlust (%), vattenhalt(%) samt GM Sand (%). Faktorn glödförlust (15-30 cm) är kvadratrots transformerad. Analysen är en Pearson’s Produkt-moment test. * anger signifikant samband (P<0,05)
Figur 8. Sambandet mellan vattenhalten på grund botten (0-15 cm) och plantornas medeldjup (cm) i 7 lokaler i
sjön Bolmen, (F1,5=14,43, P=0,02, linjär regression, R2 = 0,88).
Populationstätheten hos skaftslamkrypa i de sju inventerade lokalerna i Bolmen varierade mellan 0,35 – 2,57 plantor/m2, mätt som medel för varje lokal över hela transekt längden (0-60 cm djup, tabell 5).
Tabell 5. Medelvärde ± 95 % konfidensintervall för planttätheten mätt som medel för varje lokal över hela
transekt längden (0-60 cm)
Inventeringslokal Täthet (plantor/m2)
medel ± 95% konfidensintervall Odensjö södra 2,57 ± 0,74 Löckna 0,35 ± 0,16 Odensjö norra 1,31 ± 0,74 Hölminge 0,78 ± 0,18 Skeda 0,95 ± 0,66 Kronobäck 0,41± 0,32 Tannåker 1,07 ± 0,87 Förklaringsfaktor R2 Exponering (km) 0,41 Påväxtbiomassa (µg Chl a /mm2) 0,07 Glödförlust grunt (%) 0,01 Glödförlust djupt (%) 0,18 Vattenhalt grunt (0-15cm, %) 0,88* Vattenhalt djupt (15-30cm, %) 0,32 GMSand (%) 0,75 0,0 5,0 10,0 15,0 20,0 25,0 30,0 35,0 40,0 0,16 0,18 0,20 0,22 0,24 0,26 0,28 0,30 vattenhalt (%) medeldjup (cm)
Tätheten på plantorna kan också påverkas av de olika faktorerna. Först testades de olika lokalerna mot varandra för att säkerställa att det fanns en skillnad mellan lokaler. Detta gav att populationstätheten var skild mellan de olika lokalerna (F6,27=14,32 P<0,01, ANOVA, tabell
6).
Tabell 6. Effekten av lokal på planttätheten hos de sju inventerade lokalerna med skaftslamkrypa i Bolmen.
Analysen är en ANOVA.
Variabel Effekt SS Error SS
totalt SS DF Error Df totalt Df MS Error MS F P Planttäthet (antal plantor per m2) Lokal 16,82 5,28 22,1 6 27 33 2,80 0,20 14,32 <0,001
Odensjö Södra särskiljer sig som den lokal som tydligt har flest plantor per m2, den är även skild mot alla de övriga lokalerna (p<0,05, Tukey). Löckna och Kronobäck, med under 0,5 plantor per m2, är skilda från båda lokalerna i Odensjö(p<0,05, Tukey). Lokalerna Hölminge, Skeda och Tannåker bildar en mellangrupp som enbart är skild från Odensjö Södra (figur 9).
0 0,5 1 1,5 2 2,5 3 3,5
Odensjö södra Löckna Odensjö norra Hölminge Skeda Kronobäck Tannåker
antal plantor/m2 A B C BC BC B BC
Figur 9. Planttätheten (antal/ m2) hos 7 lokaler i Bolmen Felstaplarna anger 95% konfidensintervall. Bokstäver på staplar anger gruppering (P<0.05, Tukey)
Antal plantor per kvadratmeter var inte korrelerad med någon av faktorerna exponering, glödförlust, vattenhalt, klorofyll a eller GMSand (P>0,05, Pearson’s Produk t- moment test) (tabell 7)
Tabell 7. Korrelation mellan log x+1 planttätheten (antal plantor/m2) i de 7 inventerade lokalerna och förklaringsfaktorerna exponering (km), påväxtbiomassa(µg chl a/mm2 bladyta), glödförlust (%), vattenhalt(%) och GMSand (%).Faktorn glödförlust (15-30 cm) är kvadratrots transformerad Analysen är en Pearson’s produkt-moment test. Ingen av faktorerna har samband med planttätheten (p>0,05).
Jämförelse med lokaler utan skaftslamkrypa
Lokaler där skaftslamkrypa växte jämfördes med lokaler där skaftslamkrypa inte växte 2008. Denna undersökning visade att det fanns skillnader i sedimentets vattenhalt mellan de olika lokalgrupperna. De övriga variablerna exponering, glödförlust och GMSand visade ingen skillnad (tabell 8).
Tabell 8. Medelvärde ± 95% konfidensintervall för de olika faktorerna för lokaler med och lokaler utan
skaftslamkrypa i Bolmen. Tabellen anger även T-värde för exponering och GMSand (%) (t-test) samt Z-värde för vattenhalt och glödförlust (Mann-Withney U-test).
Förklaringsfaktor Lokaler med skaftslamkrypa medel ± 95% konfidensintervall
Lokaler utan skaftslamkrypa medel ± 95% konfidensintervall T-värde Z-värde P Exponering (km) 3,15± 1,52 2,09± 1,75 1,42 0,174 GMSand (%) 62,5± 21,7 49,6± 28,2 1,08 0,296 Vattenhalt (%) 17,0± 0,02 31,7± 0,15 -2,91 0,004 Glödförlust (%) 0,92± 0,01 3,04± 0,04 -1,31 0,191
Vattennivåns påverkan på tillväxt och reproduktion
I början av oktober insamlades plantor som vuxit barlagt och plantor som vuxit dränkt från 5 lokaler i Bolmen. För dessa mättes storlek (längd (mm), antal noder, rotlängd (mm), rotantal) och reproduktionsframgång (frön/kapsel och frön/planta, Tabell 9).
Tabell 9. Medelvärde ± 95% konfidensintervall för olika storleks och reproduktions mått hos dränkta och
barlagda plantor av skaftslamkrypa i Bolmen den 3/10 2008
Barlagda plantor Dränkta plantor
medel ± 95% konfidensintervall medel ± 95% konfidensintervall
Frön / kapsel 14,1 ± 2,8 18,4 ± 2,9 Frön / planta 34,4± 22,5 61,4± 24,2 Längd planta (mm) 25,1± 7,6 40,3± 8,3
Antal noder 8,2± 1,4 11,3± 1,8
Medel rotlängd (mm) 16,3±1,93 18,2±1,8 Medel antal rötter per nod 2,2 ± 0,2 2,8 ± 0,2
Förklaringsfaktor R2 Exponering (km) 0,48 Chl a (µg/mm2) 0,41 Glödförlust (0-15 cm %) 0,42 Glödförlust (15-30 cm %) 0,15 Vattenhalt (0-15cm, %) 0,28 Vattenhalt (15-30cm, %) 0,55 GMSand (%) 0,50
De olika storleks och reproduktionsmåtten testades mot effekten av djup och effekten av lokal. Plantor som växte dränkt hade en längre plantlängd än de plantor som växte barlagt sett till effekten av enbart djup (tabell 10) (figur 10). Det fanns däremot ingen skillnad mellan de två djupen inom lokalerna (tabell 10). Antalet noder följer samma mönster som längden på plantorna, med en effekt av djup men ingen effekt av djup och lokal (tabell 10). För antalet frön per planta finns det ingen effekt av djup men en effekt av djup×lokal(tabell 10). Antalet rötter per nod har en effekt av djupet, men denna effekt finns inte för rötternas längd (tabell 10).
Tabell 10. Effekten av djup samt djup×lokal för olika storlek och reproduktionsvariabler hos 50 plantor av
skaftslamkrypa hämtade från Bolmen den 3 oktober 2008. Analysen är en tvåvägs ANOVA.
Variabel Effekt SS Error SS totalt SS DF Error Df totalt Df MS Error MS F P Djup 0,69 8,00 1 41 0,69 1,62 0,27 Frön/kapsel Djup*lokal 1,69 7,31 9,01 4 40 44 0,42 0,18 2,31 0,07 Djup 1,71 15,82 1 41 1,71 1,80 0,25 Frön/planta Djup*lokal 3,80 14,11 17,91 4 40 44 0,95 0,35 2,69 0,04 Djup 0,65 3,119 1 41 0,65 14,87 0,02 Längd planta (mm) Djup*lokal 0,17 2,47 2,65 4 40 44 0,44 0,06 0,70 0,59 Djup 124,82 718,02 1 41 124,82 15,28 0,02 antal noder Djup*lokal 32,68 593,20 625,88 4 40 44 8,17 14,83 0,55 0,70 Djup 47,39 684,14 1 41 47,39 3,47 0,13 Medel rotlängd (mm) Djup*lokal 54,57 636,75 691,32 4 40 44 13,64 15,92 0,86 0,50 Djup 4,61 14,55 1 41 4,61 147,13 <0,01 Medel antal rötter per nod djup*lokal 0,13 9,93 10,06 4 40 44 0,03 0,25 0,13 0,97 a) b)
Figur 10. Plantlängd (mm) på skaftslamkrypa som vuxit barlagt samt dränkt. Plantorna är insamlade från 5 lokaler i
Bolmen den 3/10 2008. a) Plantlängden skiljde sig inte för de två grupperna inom lokalerna (F40,1=0,70 P=0,59,
ANOVA) ?= dränkta plantor ? =barlagda plantor. b) Det fanns däremot en effekt av enbart djup ((F40,1=14,87
P=0,0182, ANOVA). Staplar anger 95% konfidensintervall.
0 10 20 30 40 50 60 70 80 90
Odensjö södra Odensjö norra Skeda Hölminge 2 Hölminge
längd (mm) 15 20 25 30 35 40 45 50 barlagt dränkt längd (mm)
0 2 0 4 0 6 0 8 0 100 120 140 160 180 200
Odensjö södra Odensjö norra Skeda Hölminge 2 Hölminge 2
frön / planta 10 20 30 40 50 60 70 80 90 barlagt dränkt antal frö / planta a) b)
Figur 11. antal frö per planta på skaftslamkrypa som vuxit barlagt samt dränkt. Plantorna är insamlade från 5 lokaler i
Bolmen den 3/10 2008. a) Antal frön per plantalskiljde sig för de två grupperna inom lokalerna (F40,4=2,69 P=0,07,
ANOVA). Post hoc test gav dock inte svar på vart skillnaderna fanns. ?= dränkta plantor ? =barlagda plantor. b) Det fanns inte heller en effekt av enbart djup (F40,1=1,8 P=0,25, ANOVA). Staplar anger 95% konfidensintervall.
Vattennivåförhållande i Bolmen
Juni-augusti 2008 uppvisar Bolmen den för perioden lägsta vattennivån under hela mätperioden 1997-2008 vilket framgå av figur 11. Under fröbanksundersökningen i slutet av april var medel vattennivån i Bolmen 141,52 m.ö.h. Fröbanken provtogs från den nivån ner till 141,12 m.ö.h. Vattennivån hade sedan sjunkit med ca 30 cm till 141,26-141,29 m.ö.h. tills mitten av augusti då inventeringen utfördes. Under inventeringen fanns plantor på nivån 141,22 m.ö.h -140,51 m.ö.h. Vattennivåerna under 2008 innan inventeringen var som lägst 141,20 m.ö.h precis i början av augusti ,vilket framgår av figur 11. Den lägsta vattennivån för hela 2008 var den tredje oktober då medelvattennivån var 141,04 m.ö.h.
140,60 140,80 141,00 141,20 141,40 141,60 141,80 142,00
januari februari mars april maj juni juli augusti september oktober november december
vattennivå (möh)
Figur 11. Vattennivåer för sjön Bolmen 1997-2008. Gråa linjer är data för åren 1997-2007 Värden för 2008 markerade
Diskussion
Fröbanksundersökning
I denna undersökning varierade fröantalet från noll upp till 2200 frön per kvadratmeter mellan de olika lokalerna i Bolmen. För en växt som skaftslamkrypa med en kort livslängd på mindre än ett år är en hög frösättning vanlig då den enbart har möjlighet att föröka sig en gång under sin livstid (Fenner & Thompson 2005). Det här ger även möjligheten för ett stort antal frön i fröbanken som kan gro så fort förutsättningarna för dessa är de rätta (Salisbury 1967).
Bernhardt m.fl. (2004) studerade fröbanker i norra Tyskland på lokaler där skaftslamkrypa rapporterats som försvunnen. Undersökningen visade på ett stort antal frön vilande i fröbanken. Uppodling gav ca 12 000 frön medan direkt räkning gav ca 93 000 frön per kvadratmeter.
Den här studien på fröbanker i Bolmen gav en betydligt lägre fröbankstäthet än den som Bernhardts m.fl.(2004) fick genom uppodling i sin studie. Detta kan indikera att antalet vilande frön i fröbanken inte är lika hög i Sverige som i Tyskland. Ett lägre antal med frön i fröbanken innebär även färre frön som kan gro när möjlighet finns, och även mindre möjlighet att sprida sig i större antal till andra lokaler. Den stora skillnaden i resultat mellan att räkna frön direkt och att räkna dem under uppodling är viktigt att poängtera då frötätheten i denna undersökning enbart är uppskattad efter en uppodling och icke grodda frön kan förekomma.
I Bolmen var exponering den enda av de undersökta faktorerna som påverkade fröbankstätheten. Det negativa sambandet mellan exponering och antalet frön i fröbanken kan förklaras av att små frön lätt borde kunna svepas iväg av vågor och sedimentrörelser som orsakas av exponeringen. Dessa frön kan sedan sjunka till botten i lugnare vatten. Salisbury (1967) undersökte flytförmågan hos frön av skaftslamkrypa och fann att större delen av fröna sjunkit efter tre timmar i stillastående vatten. Efter totalt ett dygn flöt fortfarande tio % av fröerna. En fördröjning av att fröerna sjunker betyder att fröerna kan spridas över en större yta och ger möjlighet för växten att etablera sig på nya lämpliga lokaler. Det här innebär även att de kan spridas till habitat som är olämpliga, till exempel upp på land eller till för djupt vatten eller till lugnare vikar där frön av skaftslamkrypa riskerar att försvinna i djupare
sedimentlager. Att fröer flyter på ytan behöver inte heller betyda att de lätt flyter upp till ytan om de inte påverkas av rörelser i vattnet
Lutningen på det linjära sambandet mellan exponering och frötätheten kan komma att ändras om fler lokalers fröbanker undersöks då referenslokalen som slumpades ut hade en avvikande hög exponering. Då antalet lokaler enbart var fem var det också svårt att få en normalfördelning på data som behövs för att testa korrelationer. Fler datapunkter hade troligen gett en bättre normalfördelning och ett tydligare resultat.
Djuputbredning och täthet
Djupet som skaftslamkrypa växte på varierade mellan 3-74 cm djup under inventeringen i augusti. Den var skild från bottnens medeldjup på två lokaler av sju vilket innebär att växtdjupet på dessa lokaler inte är slumpmässig utan troligen styrd av någon faktor. Tätheten på plantorna varierade mellan 0,35 – 2,57 plantor/m2, mätt som medel för varje lokal över hela transekt längden (0-60 cm).
Av faktorerna exponering, glödförlust, vattenhalt och GMSand var det enbart sedimentets vattenhalt på grunt vatten (0-15 cm) som visade ett samband med medeldjupet på plantorna (figur 8). Vattenhalten är ett mått på bottnens hårdhet men även hur finkornig den är då finare sediment kan binda mer vatten till sig. I lösare sediment ökar risken för att frön begravs för djupt för att kunna gro, då ljus kan vara en avgörande faktor för att groning ska starta (Salisbury 1967, Fenner och Thompson 2005). På grundare vatten har vågor även en mer direkt verkan på botten än på djupare vatten vilket kan störa etablering av plantor (Koch 2001). Lösare sediment på grundare vatten kan öka denna effekt då plantor saknar möjlighet at få fäste i sedimentet.
Undersökningen av mängden påväxtbiomassa (µg klorofyll a per mm2 bladyta) på blad av notblomster gav ingen koppling till djuputbredningen hos skaftslamkrypa. Påväxter kan tänkas påverka djuputbredningen då de bidrar till skuggning i en miljö där ljustillgången redan kan vara begränsad. Halten klorofyll a på notblomster varierade mellan 1,12-7,68 µg chl a /mm2. Detta kan jämföras med en studie av Weisner m.fl. (1997) där mängden påväxter i två eutrofa sjöar i Sverige undersöktes för skyddade samt exponerade lokaler. I den studien varierade mängden påväxter mellan ca 10µg chl a /mm2 till ca 90 µg chl a /mm2 där de högre värdet representerar skyddade lokaler. Dessa halter är tio gånger högre än halterna i Bolmen.
Låga halter av klorofyll a kanske därför kan förklara varför påväxter inte påverkar utbredningen av skaftslamkrypa i Bolmen.
Odensjö södra är den lokal från inventeringen 2006 som har lägst antal frön i fröbanken (<1000 frön/m2) men den är också den lokal som har den högsta organiska halten inom fröbanksintervallet (ca 35 %, bilaga 3 tabell 3).Här hittades också flest plantor under inventeringen (N= 395), men den organiska halten var vid dessa plantor betydligt lägre (ca 1 %). Anledningen till detta mönster kan vara att skaftslamkrypa enbart växer i sediment som har en lägre organisk halt där plantor inte blir utkonkurrerade. Något samband till den organiska halten upptäcktes inte i denna studie, men som diskuterats tidigare behövs troligen fler lokaler för att ett eventuellt mönster ska framträda.
Att samband saknas för skaftslamkrypa i Bolmen kan även bero på att lokaler med skaftslamkrypa som till synes inte är lämpliga som habitat kan inhysa mindre områden med bättre förutsättningar (Grime 2001). Bolmens stränder är blockrika och det finns gott om stenar, både stora och små, som kan ge bra skydd mot en direkt exponering under växtsäsongen. Under inventeringen märktes detta tydligt genom att flertalet av lokalerna hade vass eller sjösäv och även stenar eller uppgrundningar i bottnen där skaftslamkrypa växte skyddat bakom. I hårt exponerade lokaler där bottnen nästan enbart består av klappersten kan skaftslamkrypa fortfarande växa om den bara hittar fäste i sedimentet mellan stenarna där den kan växa relativt skyddat (se figur1 bilaga 2).
I denna undersökning påträffades inga plantor på grundare vatten som barlagts tidigt under växtsäsongen (juni-juli). Förklaringen till detta mönster kan vara ljuskonkurrens från andra växter. I Bolmen växte skaftslamkrypa tillsammans med andra karaktäristiska arter för oligotrofa sjöar som till exempel notblomster, nålsäv (Eleocharis acicularis) och strandpryl (Litorella uniflora). Tätare mattor med dessa växter, som förekom främst på grundare vatten, verkar kunna hindra kolonisation av skaftslamkrypa, då denna främst växte i den yttre periferin av dessa mattor. Då de ovan nämnda arterna är perenna och redan etablerade inför nästkommande växtsäsong har de en fördel jämfört med skaftslamkrypa som troligen är annuell och måste återetablera sig varje år.
Skaftslamkrypa växte i Bolmen även inne i glesa bestånd av bladvass och sjösäv men observerades inte i täta bestånd som skuggade mycket. Observationerna från denna
undersökning överensstämmer med studien av Bernhardt m.fl. (2008). Här växte skaftslamkrypa främst i den zonen som karaktäriserades av pionjärväxter, i det område där botten sällan blev barlagd, vilket möjliggjorde för öppna ytor där skaftslamkrypa kunde etablera sig. Den förekom även i samma zon som sjöfräken (Equisetum fluviatile) då skaftslamkrypa kunde växa i de luckor som bildades (Bernhardt m.fl. 2008).
Resultatet från undersökninge n av växternas djupfördelning visar att lokaler i sydvästra delen av Bolmen (lokalerna i Odensjö och Löckna) har ett plantmedeldjup som är grundare än i lokalerna i nordöst (Skeda, Kronobäck och Tannåker) (tabell 2). Bolmen uppvisar tack vare sin storlek och utformning skillnader i vattenkemi mellan norra och södra delen av sjön (Hein 1999). Dessa olika faktorer skulle kanske kunna förklara varför plantdjupet är skiljt från medeldjupet på lokalerna.
Jämförelse med lokaler utan skaftslamkrypa
I lokaler där skaftslamkrypa inte växte var vattenhalten högre än i lokaler med skaftslamkrypa. Lösare bottnar kan innebära svårigheter för skaftslamkrypa genom fasthållningsproblem och av att bladen skuggas av sediment, vilket diskuterats tidigare. Det mönster som framträder, men inte är fastställt i undersökningen, är en tendens till högre exponering och en större andel grövre sediment i lokaler med skaftslamkrypa mot mer organiska lösare bottnar i lokaler där de inte förekommer. Detta stöds inte av undersökningens resultat då enbart vattenhalten var skild mellan de lokaler där skaftslamkrypa fanns och referenslokaler. En undersökning med fler data än vad som användes i denna undersökning skulle behövas för att fastställa om fler faktorer kan kopplas till förekomst av skaftslamkrypa.
Vattennivåns påverkan på tillväxt och reproduktion
En planta har begränsat med resurser under sin livstid som den ska fördela på bland annat tillväxt och reproduktion men andelen resurser som läggs på reproduktion är oftast fixerad inom arten (Fenner och Thompson 2005). Detta skulle innebära att större plantor kan producera en större andel frön än plantor av en mindre storlek. Skaftslamkrypa som växte dränkt i Bolmen var generellt längre och hade fler noder än de barlagda plantorna (tabell 9). Att plantorna blir längre under vatten kan vara en kompensation för de lägre ljusförhållanden som råder under vattenytan då en mer utbredd fotosyntetiserande yta klarar av att ta upp mer ljus och därmed bibehålla sin fotosyntetiserande förmåga. Detta ger även en större yta för gasutbyte då nivåerna av koldioxid och syre är lägre i vatten än i luft (Sculthorpe 1985).
Hos plantor i Bolmen var antal frön per kapsel 18,4 för dränkta plantor och 14,1 för barlagda plantor (tabell 9). I Salisburys (1967) odlings försök med dränkta samt barlagda plantor var motsvarande antal 16,6 respektive 30,0 frön per kapsel. Antalet frön per kapsel i denna studie skiljer sig inte mellan de båda djupen, vilket inte stödjer mönstret i Salisburys (1967)studie Fröantalet per kapsel hos plantor från Bolmen skiljer sig inte från fröantalet för de dränkta plantorna i Salisburys (1967) undersökning. En förklaring kan vara att plantorna i Bolmen delvis varit dränkta och kanske även blommat under vatten vilket gör att fröantalet/kapsel blir samma som för plantor som varit konstant dränkta då inga blommor utvecklats som kan korspollineras.
Fröproduktionen per planta visade inte heller någon effekt av vattendjupet, denna effekt fanns troligen enbart inom vissa lokaler. Detta innebär att även om plantorna är längre och har fler noder när de växer dränkt finns det ingen koppling till en ökad fröproduktionen per planta, en större storlek under vatten ger inte en högre reproduktiv förmåga. Medel fröproduktionen per planta, bortsett från växtdjupet, var i Bolmen ca 50 frön per planta. I Salisburys studie var samma värde för barlagda plantor ca 6700. Detta innebär att antalet frön som produceras per planta är betydligt lägre i Sverige, vilket även förklarar det lägre antalet frön i fröbanken.
En väsentlig faktor för reproduktionsframgången är även kvaliteten på fröna som plantorna producerar vilket inte undersöktes i denna studie. Kvaliteten mellan frön producerade av barlagda plantor kan skilja sig från kvaliteten på frön från dränkta plantor, speciellt då de från dränkta plantor främst kommer från självpollinerade blommor.
Vattennivåförhållande i Bolmen
Skaftslamkrypa som hittades under inventeringen i den andra delen av augusti växte alla under vatten. Plantor på grundare vatten (0-10 cm) hade tidigare vuxit barlagt då vattennivån i början av augusti var ca 5 cm lägre än referensnivån 141,25 m.ö.h. Denna barläggning varade som längst en vecka. När vattennivån började sjunka under slutet av augusti ner till den lägsta nivån 141,05 m.ö.h. kom plantor som tidigare vuxit ner till 20 cm djup att bli torrlagda. Plantor under denna nivå växte enbart under vatten under 2008.
Av dessa plantor hade enbart ett fåtal vuxit barlagt tidigare under säsongen. Den lägsta vattennivån innan inventeringen var 141,20 m.ö.h. (3/8) och plantorna var barlagda som
längst en vecka. Detta gällde dessutom enbart lokalerna i Odensjö, då de övriga lokalerna som inventerades inte hade plantor som växte så grunt. Den lägsta vattennivån 2008 var den 3/10 då nivån var 141,04 m.ö.h. Detta innebär att plantor som växte djupare än ca 20 cm aldrig kom att växa barlagt. Detta stämmer inte överens med den övriga litteraturen då arten i Danmark, Nederländerna respektive England beskrivs främst som växande på land och mycket sällan i vatten (Salisbury 1967, Thyssen 1990, Brinkkemper m.fl. 2008). Salisbury (1967) nämner att skaftslamkrypa har hittats växande under vatten både i England och utomlands, troligen från frön som grott barlagt men åter blivit dränkta.
Anledningen till att skaftslamkrypa inte växer barlagt i större utsträckning kan vara, som tidigare nämnts, en högre konkurrens längre upp på stranden där växtligheten är tätare. Även klimat faktorer kan tänkas påverka då skaftslamkrypa i Sverige växer på sin nordligaste gräns vilket utsätter groddar och plantor för en större variation i temperatur som inte blir lika påtalig under vatten.
Slutsatser och idéer för bevarande av skaftslamkrypa
Antalet vilande frön i fröbanken var i denna studie lägre än vad som funnits i Tyskland av Bernhardt m.fl. (2004). För bevarande av skaftslamkrypa är det viktigt att man kan fastställa om den finns i fröbanken, och i vilket antal. Detta gäller både på lokaler där den förekommer men även på lokaler där den förekommit tidigare då den fortfarande kan finnas vilande i fröbankerna. Detta skulle kunna innebära att man genom åtgärder för att återställa en lämplig miljö för arten, som till exempel rätt vattenkvalitet och mindre skuggning från annan vegetation, kanske kan aktivera de frön som redan finns i sedimentet.
Lämpliga lokaler för skaftslamkrypa verkar vara lokaler som inte utsätts för en direkt exponering under växtsäsongen men ändå har en relativt fast botten. Bottnar i lugnare lokaler med lösare sediment är troligen olämpliga då plantor och frön antingen begravs i sedimentet eller utkonkurreras av andra arter.
I en reglerad sjö är det även viktigt att fastställa på vilken nivå som fröbankerna finns, då skaftslamkrypa verkar växa företrädesvis under vatten. Preferenser för groning under vatten eller inte återstår att besvara, detta gäller även hur bra de i så fall klarar sig efter groning. Även tidpunkten på året då groning sker är intressant att undersöka, då detta tillsammans med kunskapen om vart de gror (barlagt/dränkt) kan kopplas till vattenregimen för respektive sjö.
Kopplingen till de olika faktorerna var svag i denna studie. Detta kan bero på olika förklaringar, till exempel lokalernas utformning som diskuterats innan, men även att antalet prov (5-13 per undersökning) var för få för att ett eventuellt samband ska framträda. Ytterligare undersökningar med fler provpunkter skulle kunna ge en tydligare bild av förhållandet mellan skaftslamkrypa och de olika faktorerna.
Det finns indikationer på att skaftslamkrypa ökat i Nederländerna under 1900-talet (Brinkkemper m.fl. 2008). De nämner att arter inom släktet Elatine har ett stort
utbredningsområde, men inom det är den fraktionerad i tid och rum. I Sverige finns för få undersökningar för att ge en korrekt bild av förekomst och utbredning(Artdatabanken 2007). Om det är en naturlig fluktuation i populationen eller en signal på att den är på tillbakagång kan därmed vara svårt att besvara. Brist på studier tillsammans med observationssvårigheter möjliggör att den idag kan vara mer förekommande än man tidigare trott. Fler inventeringar av skaftslamkrypa behövs för att fastställa hur den klarar sig i Sverige och hur spridd den egentligen är, både som plantor och i fröbank.
Tack
Börje Ekstam, Högskolan i Kalmar Marie -José Guillard, Högskolan i Kalmar Catherine Legrand, Högskolan i Kalmar Per Ekerholm Länstyrelsen i Kronoberg Krister Wahlström, Skogstyrelsen, Kjell Wiktorsson, EON
Sara Gunnarson och Agneta Häggerud Högskolan i Kalmar
Släkt och vänner, för all support, stöd och handräckning, detta arbete hade inte tillkommit utan er Till alla markägare kring Bolmen och till Bolmens camping ett extra tack för all er gästfrihet
Referenser
Artdatabanken. Bertilsson A, 1995. Faktablad; Elatine hexandra – skaftslamkrypa, http://www.artdatabanken.se, 2007-01-04.
Bernhardt K.-G., Koch M., Kropf M., Ulbel E., Webhofer J, 2008. Comparison of two methods characterising the seed bank of amphibious plants in submerge d sediments. Aquatic botany 88: 171-177.
Bernhardt K.-G., Koch M., Kropf M., Ulbel E., Webhofer J., 2004. The soil seedbank as a resource for in situ and ex situ conservation of extinct specie s., Scripta Bot. Belg.,29: 135-139.
Brinkkemper, O., Weeda E.J., Bohncke S.J.P. Kuijper W.J., 2008. The past and present
occurrence of Elatine and implications for palaeoenvironmental reconstructions, Veget Hist Archaeobot 17: 15-24.
Fenner M. , Thompson K., 2005. The ecology of seeds. Cambridge University Press, Cambridge Grime J.P, 2001. Plant strategies, vegetation processes and ecosystem properties, second edition.
John Wiley & Sons Ltd, New York.
Gärdenfors, U. (ed.), 2005. Rödlistade arter i Sverige 2005 – The 2005 Red List of Swedish Species. ArtDatabanken, SLU, Uppsala.
Hein M. (red.), 1999. Bolmen 2000 vattenkvalitet En sammanställning av fysikaliska, kemiska och biologiska parametrar i sjön Bolmen 1966-1999. Länsstyrelsen i Jönköpings län
meddelande nummer 1999: 48.
Hutchinson, G. E., 1975. A treatise on limnology, Volume III, Limnological botany. John Wiley & Sons, New York.
Jespersen A-M. , Christoffersen K., 1987. Measurments of chlorophyll-a from phytoplankton using ethanol as extraction solvent. Arch. Hydrobiol. 109: 445-454.
Keddy, P.A., 1982. Quantifying within-lake gradients of wave energy: interrelationship of wave energy, substrate, particle size and shoreline plants in Axe Lake, Ontario. Aquatic Botany 14: 41-55.
Koch E.W., 2001. Beyond light: physical, geological, and geochemical parameters as possible submersed aquatic vegetation habitat requirements. Estuaries 24: 11-17.
Madsen J.D., Chambers P.A, James W.F., Koch E.W., Westlake D.F., 2001. The interaction between water movement, sediment dynamics and submersed macrophytes. Hydrobiologia 444: 71-84.
Naturvårdsverket, 2003. Natura 2000 Art- och naturtypsvisa vägledningar, vattenmiljöer och gräsmarker 1. Http://www.naturvardsverket.se/, 2008-06-10.
Rasband W.S., 2008. ImageJ. U. S. National Institutes of Health, Bethesda, Maryland, USA.
Http://rsb.info.nih.gov/ij/, 2008-09-09.
Salisbury E. J.,1967. On the reproduction and biology of Elatine hexandra; a typical species of exposed mud. Kew Bulletin , 21: 1 139-149.
Sculthorpe C.D., 1985. The biology of Aquatic Vascular Plants. Edward Arnold Publishers, Königstein.
SMHI. Vinddata för Torups väderstation. Http://www.smhi.se,2008-07-14.
Svensk standard, SS 02 81 13. Vattenundersökningar – bestämning av torrsubstans och glödgningsrest i vatten, slam och sediment.
Ter Heerdt G.N.J, Verweij G.L., Bekker R.M. , Bakker J.P., 1996. An improved method for seed-bank analysis: seedling emergence after removing the soil by sieving. Functional Ecology 10: 144-151.
Thompson K., Bakker J.P., Bakker R.M., 1997. The soil seed banks of north west Europe: methodology, density and longevity. Cambridge University press, Cambridge.
Thyssen N. (red.), Miljøministeriet miljøstyrelsen 1990. Danmarks miljøundersøgelser; Danska vandplanter vejledning; bestemmelse af planter i søer og vandløb; Bækarve (Elatine) og Vandportulak (Peplis). Miljønytt nr 2.
Wahlström K. (observatör). Rapportsystemet för växter och svampar; Portal för växter, mossor, svampar, lavar och alger. Http://www.artportalen .se, 2008-04-15.
Weisner S.E.B., 1987. The relation between wave exposure and distribution of emergent vegetation in a eutrophic lake. Freshwater biology, 18: 537-544.
Weisner S.E.B. Strand J.A., Sandsten H., 1997. Mechanism regulating abundance of submerged vegetation in shallow eutrophic lakes. Oecologia 109: 592-599.
Willby N.J., Abernethy V.J., Demars B.O.L., 2000. Attribute-based classification of European hydrophytes and its relationship to habitat utilization. Freshwater Biology, 43: 43-74.
Bilaga 1) Kartor
Figur 1. Karta med koordinater för de lokaler som inventerades i augusti 2008; lokal A saknar
koordinater men denna var belägen ca 50 m söder om lokal B. Lokaler 1-6 användes enbart till förekomst undersökningen. Lokal X Y 6306897 1367044 B) Odensjö norra 6306939 1367043 6310176 1368371 C) Löckna 6310204 1368370 6311609 1376476 D) Hölminge 6311617 1376455 6315995 1376489 E) Tannåker 6315993 1376501 6321435 1379486 G) Kronobäck 6321417 1379503 6321034 1376684 F) Skeda 6321060 1376702 6316027 1376479 1) Tannåker östra 6316028 1376485 6315693 1375867 2)Tannåker västra 6315701 1375883 6311621 1376415 3)Hölminge västra 6311626 1376438 6300639 1366334 4) Inre jättabron 6300667 1366352 6300972 1366853 5) Mittersta jättabron 6300977 1366839 6301029 1366982 6) Yttre jättabron 6301021 1366946
Figur 2. Karta med koordinater för referenslokaler i Bolmen där skaftslamkrypa inte hade någon
förekomst 2008
Figur 3. Karta med koordinater för lokaler från 2006 som användes till fröbanksundersökningen
(data från Krister Wahlström), samt en referenslokal där skaftslamkrypa inte hade någon förekomst 2006. Lokal X Y 1 6302647 1365710 2 6304001 1365551 3 6307808 1367437 4 6318438 1375569 5 6318898 1375292 6 6320917 1376453 7 6322319 1377324 lokal X Y 1 Skällandsö 6320657 1377824 2 Tannåker 6315735 1375520 3 Bolmstad 6308450 1375857 4 Jättabron 6300802 1366566 5 Odensjö 6366807 1367031
Bilaga 2) Foton
figur 1. Skaftslamkrypa (inringad) växande bland stenar på botten av sjön Bolmen tillsammans med notblomster
(Lobelia dortmanna) och strandpryl (Litorella uniflora). Foto: A . Edlund ©
a) b)
figur 2. närbild av skaftslamkrypa i en av burkarna i fröbanksförsöket, längd mellan stora bladen 3 mm 2b) alla burkar
i försöket (foto: A. Edlund 25/6 2008)
Figur 3. Skaftslamkrypa (inringad) samt förväxlingsart (notblomster)i fröbanksundersökningen. Märk att
förväxlingsarten enbart bildar ett långt blad efter hjärtbladen medan skaftslamkrypa forsätter vara parbladig. (Foto: A. Edlund 25/6 2008)
a b
Figur 4. a)Frön av skaftslamkrypa där den mörka, översta, är ett moget frö. Längd på frö ca 2mm (Foto: A. Edlund) b) Skaftslamkrypa med blomma uppodlad under fröbanksundersökningen .
4a 4b
Figur 5. Två typiska stränder vid Bolmen. Bilder från inventeringslokal a) Kronobäck i norra delen av Bolmen samt b) Odensjö södra i södra delen. Foton tagna i samband med inventeringen i augusti 2008. (Foto: A. Edlund)